Vous venez de mettre les pieds dans les tréfonds en construction de ControverSciences.
Simplification adaptative et évolution de la locomotion chez le gecko : conséquences morphologiques et bio-mécaniques de la perte d'adhésion
Adaptive simplification and the evolution of gecko locomotion: Morphological and biomechanical consequences of losing adhesion
Timothy E. Higham, Aleksandra V. Birn-Jeffery, Clint E. Collins, C. Darrin Hulsey, Anthony P. Russell
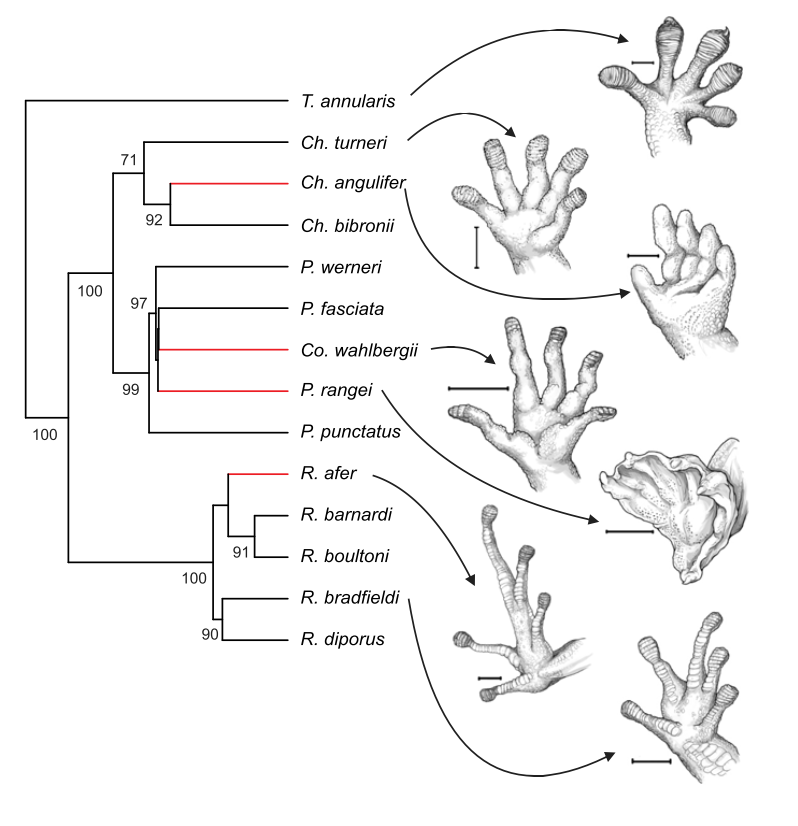
FIg. 1. Relations phylogénétiques parmi les 14 espèces de Gecko examinées dans cette étude, associées à l'illustration de la partie ventrale du pied arrière de 7 d'entre eux. Timothy E. Higham, Aleksandra V. Birn-Jeffery, Clint E. Collins, C. Darrin Hulsey, Anthony P. Russell, 2014
Les lignées évolutives se sont diversifiées au cours du temps par la mise en place d'innovations. Ces innovations peuvent néanmoins aussi imposer des contraintes fonctionnelles. C'est pourquoi, lorsque les pressions de sélection se relâchent, les contraintes diminuent, et des simplifications secondaires peuvent potentiellement avoir lieu.
La locomotion du gecko et les adaptations morphologiques et bio-mécaniques qui y sont liées sont un bon exemple de cette éventuelle simplification secondaire. En effet, suite à des pressions de prédation fortes, le gecko est devenu un excellent grimpeur en développant un système adhésif exceptionnel. Le retour à un mode de vie terrestre pourrait donc impliquer une perte de ces fonctions et caractéristiques hors du commun.
Le genre Pachydactylus est le genre le plus documenté et donc le plus facile à analyser.
L'expérience a consisté en la comparaison des conséquences de la réduction et de la simplification de l'appareil parmi 14 espèces du genre Pachydactylus (dont deux espèces - Chondrodactylus angulifer and Pachydactylus rangei - montrent des pertes du système adhésif évolutivement indépendantes et non ambigues, et deux espèces -Rhoptropus afer and Colopus wahlbergii- présentent des réductions significatives) afin de tester si les espèces présentant ces réductions:
i) présentent aussi des cinétiques des membres différentes
ii) présentent des taux plus élevés d'évolution de la cinétique et de la morphologie
iii) présentent différentes réponses cinétiques aux changements d'inclinaison du substrat
iv) courent plus vite ou plus lentement que celles qui ont un système adhésif complet.
Les analyses statistiques consistent en des ACP
La réduction ou la simplification du système adhésif modifie de manière significative la manière qu'ont les geckos de courir. En plus de changement dans la vitesse et la cinétique du mouvement, le taux d'évolution de la morphologie est plus important chez les espèces qui démontrent une perte ou une simplification de leur appareil adhésif. Ce qui suggère que les contraintes associées à l'adhésion ont été contournées. L'association entre un mode de vie terrestre et la perte/réduction de l'adhésion suggère un lien direct entre la morphologie, la bio-mécanique et l'écologie. Des études antérieures ont montré l'existence de compromis évolutifs entre les capacité de sprint et grimpe des geckos. Il n'est donc pas surprenant que les geckos terrestres subissent des réductions ou simplifications évolutives de leur système adhésif.
Donc non seulement une perte de traits morphologiques est possible au cours de l'évolution, en conséquence à un relâchement de pression de sélection, mais en plus elle peut s'accompagner de simplification de mouvements, afin de satisfaire aux compromis évolutifs mis en jeu.
Dernière modification il y a plus de 10 ans.
Plant interactions with microbes and insects : from molecular mechanisms to ecology
Plant interactions with microbes and insects: from molecular mechanisms to ecology
Corné M.J. Pieterse, Marcel Dicke
Les plantes sont en interaction constante avec une multitude d’organismes qu’ils soient bénéfiques ou non. Les associations mutualistes permettent à la plante de résister à des stress environnementaux. Une reconnaissance mutuelle et une communication est nécessaire. Les plantes doivent faire face à des invasions de pathogènes et d’herbivores et donc développer et accumuler au cours de leur évolution des stratégies sophistiquées pour percevoir les interactions biotiques et y répondre comme il se doit.
La première ligne de défense, est la reconnaissance des caractéristiques communes des pathogènes, il s’agit du niveau de base de la résistance. Au cours de la coévolution, les pathogènes ont acquis la capacité de supprimer cette défense, et les plantes se sont ensuite complexifiées pour contrer cette attaque par l’acquisition de nouvelles défenses, une deuxième ligne de défense apparaît, l’immunité effectrice-déclencheur. La résistance qui agit dans toute la plante, s’ajoute aux défenses immunitaires déclenchées par reconnaissance des attaquants. Ces mécanismes impliquent la production de composés défensifs et la diffusion d’hormones spécifiques qui permettent de réguler les réactions de défense suivant le type d’attaquant, l’attaque ou le bénéfice.
Les plantes interagissent avec une multitude d’organismes ce qui rend ses capacités très complexe à répondre à son environnement biotique et un équilibre entre protection et bénéfice est appliqué. Ainsi un chevauchement complexe des réseaux de signalisations activés par les parasites, herbivores ou mutualistes apparaît. De plus la communauté complexe et les caractéristiques individuelles sont à l’origine d’une variation intraspécifique.
Les signaux de défenses varient en fonction du type d’organisme qui interagit. La quantité, la composition et le temps du signal provoque l’activation d’un ensemble de gènes spécifique à la combinaison plante-attaquant, pour répondre efficacement et sans perte d’énergie à l’interaction. Il est aussi apparu d’autres mécanismes régulateurs qui forment une réponse défensive complexe, comme la diaphonie ou les signaux induits par l’attaquant. Les voies hormonales interagissent entre elles et leur activation simultanée peut avoir des effets positifs (amélioration des défenses) ou négatifs (antagonisme). De plus différentes protéines régulatrices ont un rôle dans les défenses et sont des facteurs de transcription de certains gènes de défense. L’apparition de la diaphonie entre les voies de défenses permet à la plante de réagir efficacement aux attaques, et de choisir la stratégie à suivre en fonction du type d’attaquant.
Cependant, les organismes qui interagissent avec les plantes ont aussi évolué vers une manipulation de cette dernière pour leurs propres bénéfices, en acquérant des mécanismes bloquant les défenses. Certains pathogènes suppriment ces défenses par le biais de facteurs de virulences. De plus la variabilité intraspécifique dans la façon dont les voies de défense communiquent chez les plantes, permettrait de contrecarrer les réponses des attaquants.
Un niveau de complexité s’ajoute à la manière qu’ont les plantes de s’adapter à leur environnement biotique : l’amorçage pour la défense renforcée. Les défenses ne sont pas activées directement par les attaques, mais sont plus ou moins accélérées suivant la perception des signaux de stress. Les coûts sur la fitness des plantes semblent plus faibles que ceux induits par les défenses directes. Ces fonctions d’amorçage sont une adaptation qu’ont les plantes à répondre rapidement à leur environnement. L’émission de volatiles spécifiques à l’herbivore va permettre aux plantes voisines de réagir rapidement en améliorant leurs défenses directes et indirectes. Ceci est un avantage écologique pour les plantes qui vont produire en réponse à des signaux extérieurs les défenses nécessaires, leur permettant d’éviter la dépense d’énergie dans la fabrication continuelle de défenses.
Les traits de défense sont bien souvent polygéniques et varient fortement ce qui forme un « réservoir » de mécanismes d’adaptation qui ont évolué pour répondre aux stress biotiques. Il semblerait que ce niveau élevé de complexité atteint par la plante pour répondre efficacement à son environnement biotique serait le produit de la course à l’armement de la coévolution entre les plantes et leurs agresseurs. Pour mieux comprendre le fonctionnement des réseaux de signalisations complexes de défense, une approche moléculaire et écologiste est nécessaire afin de placer les mécanismes moléculaires des défenses dans une perspective écologique.
La course à l'armement apporte et accumule de nouveaux moyens de défense permettant à la plante de contrecarrer les attaques des pathogènes et herbivores. On a pu voir ici à l'échelle individuelle que les mécanismes de défense se complexifient avec la coévolution.
Dernière modification il y a plus de 10 ans.
The origins of genome complexity
The Origins of Genome Complexity
Michael Lynch and John S. Conery
Il est observé au cours de l’évolution des eucaryotes un développement de la taille et de la complexification des génomes avec une augmentation du nombre de gènes, une apparition de nouvelles fonctions, une augmentation du nombre d’introns et une prolifération des éléments génétiques mobiles.
De plus il est important de noter une réduction de la taille des populations lors de la transition de procaryote à eucaryote unicellulaire puis à eucaryote multicellulaire. Une taille de population plus petite engendre une dérive génétique plus forte, et donc une prolifération de diverses caractéristiques génomiques qui seraient éliminées par sélection purificatrice chez les populations de grandes tailles. Ainsi la force de la dérive génétique semblent varier fortement entre les organismes unicellulaires les plus petits et les multicellulaires les plus grands. La taille efficace de la population et le taux de mutation varient fortement entre procaryote, eucaryote unicellulaire, plante, invertébrés et vertébrés.
Il semblerait que l’augmentation du nombre de gènes dans les populations soit due à des processus stochastiques d’apparition et de perte de gènes. Les gènes nouvellement dupliqués peuvent être perdu suite à l’accumulation de mutations délétères plus susceptible d’apparaître. Mais on peut voir la préservation des deux membres d’une duplication, qui sera favorisée par l’apparition d’une mutation bénéfique sur un membre modifiant la fonction d’origine conservée par l’autre duplicat, ainsi cette néo-fonctionnalisation va permettre l’apparition d’un nouveau gène. De plus suite à une duplication d’un gène à fonctions multiples, les deux membres du couple peuvent être partiellement dégradés favorisant l’obtention de multiples gènes avec des fonctions complémentaires. Cette sous fonctionnalisation va permettre l’élimination des contraintes pléiotropiques des gènes ancestraux. Ces deux phénomènes de néo- et de sous-fonctionnalisation dépendent fortement de la taille de la population. Les populations de petite taille conserveraient plus longtemps les gènes nouvellement dupliqués qui seront sujets à la sous-fonctionnalisation. Ainsi l’évolution de la multicellularité serait due non seulement aux nouveaux défis sélectifs contrés grâce à la néo-fonctionnalisation mais également une partie de l’augmentation du nombre de gène peut ne pas avoir été conduit par des processus adaptatifs mais plutôt suite à une réponse passive à un environnement génétique (taille de population réduite) plus propice à la préservation des gènes dupliqué par sous fonctionnalisation
De plus une augmentation de la quantité et de la taille des introns est observable avec l’augmentation de la taille du génome. Seulement ceci ne semble pas présenter d’avantage évident. Les introns nouvellement établis peuvent imposer un désavantage sélectif sur leur gène en augmentant le taux de mutation mais aussi la présence de la région d’épissage sur ces introns est indispensable pour la traduction. Les mutations peuvent affecter cet épissage et le bon déroulement de l’expression des gènes. Ainsi la prolifération des introns dépendra du taux de mutations et de la taille du génome. Les introns peuvent permettre également un épissage alternatif augmentant la possibilité d’action des gènes. Présents dans la plupart des gènes, les introns subissent un mécanisme fiable pour l’épissage et ils fournissent un mécanisme d’orientation pour la surveillance des ARNm défectueux.
Les éléments génétiques mobiles sont des unités génomiques indépendantes capables de se fixer sur le génome. Ils peuvent s’insérer n’importe où dans le génome et peuvent le modifier. Les mutations avec des conséquences négatives sur la fitness sont efficacement éliminées par la sélection. Ils peuvent cependant apporter un certain bénéfice à l’organisme et être conservés. La fixation des éléments mobiles dans le génome va dépendre fortement de sa taille, et de celle de la population.
La taille de la population et les mutations semblent guider l’évolution génomiques des diverses lignées phylogénétiques. L’évolution du génome des organismes serait due à des réponses indirectes de la réduction de la taille des populations.
Seules les modifications bénéfiques dans le génome seront conservées au cours de l’évolution, l'ajout d’intron et d’éléments mobiles n’apportant pas de désavantages sélectifs, la néo- et sous-fonctionnalisation qui apportent de nouveaux gènes et de nouvelles fonctions suite à la duplication. Ainsi on pourrait sous entendre que l’évolution du génome entraîne une complexification pour l’ajout de nouvelles fonctions dû à des mutations ou des duplications par exemple. D'autres moyens peuvent générer de multiple fonctions de gènes, comme la régulation de gène tissus spécifique (conséquence de la sous-fonctionnalisation), ou l'épissage alternatif (fixation d'intron).
De ce fait l'évolution du génome pourrait conduire à sa complexification.
Dernière modification il y a plus de 10 ans.
The complexity and stability of ecosystems
The complexity and stability of ecosystems
Stuart L. Pimm
La richesse spécifique, les interactions et l’uniformité du système définissent la complexité. Un écosystème est dit stable s'il revient à l’équilibre initial après une perturbation. Les hommes ont tendance à simplifier les environnements naturels rendant les écosystèmes moins stables. Ceci pourrait être la conséquence d’une augmentation des populations instables conduisant à l’extinction de certaines puis à la simplification. Ceci semble établi, cependant existe-t-il réellement une relation entre stabilité et complexité ?
Des études montrent que l’évolution des écosystèmes vers un système plus complexe améliore leur stabilité. Ceci semblait si certain que l’idée est devenue centrale. Mais c'est un sujet très controversé, quand certains montrent que l’accumulation de la diversité biologique favoriserait la stabilité des communautés, d’autres présentent le contraire. La théorie et l’expérimentation présentent des résultats hétérogènes et incomplets, soulignant l’intérêt de l’étude de cette relation.
Les différents modèles de simulation de la dynamique des systèmes sont eux même plus ou moins complexe et prennent en compte des paramètres plus ou moins pertinents et représentatifs de la réalité.
Les systèmes agricoles par exemple, pauvres en espèces donc peu complexes montrent des risques accrus face aux ravageurs contrairement aux systèmes tropicaux très diversifiés.
Les organismes vont plus facilement coloniser des îles pauvres en espèces, cette simplicité montre la facilité à s’installer. Les modèles suggèrent que les communautés avec plus d’espèces sont plus fortes face à l’envahissement, moins il y a d’espèces dans la communauté plus la probabilité d’arrivé est élevée, ainsi les écosystèmes avec peu d’espèce sont susceptibles d’évoluer vers une complexification en augmentant leur richesse spécifique. Le système est donc modifié par l’apport de nouvelles espèces qui vont changer l’équilibre, ainsi un système simple sera susceptible de se complexifier par l’ajout de nouvelles espèces.
Les interactions entre les espèces vont également s’ajouter et complexifier le système. Les interactions interspécifiques peuvent engendrer une certaine complexité du système même avec peu d’espèces, le rendant plus résistant et stable. Des modèles montrent que l’augmentation de la connectance et de la richesse spécifique va augmenter la stabilité de l’écosystème, plus le système est complexe plus il est stable. Cependant certains modèles montrent que des systèmes trophiques complexes sont plus fragiles et sujets aux extinctions que les systèmes simples. De plus des systèmes simples persisteraient plus longtemps que des complexes, posant la question : est ce que l’abondance d’espèces des communautés de désert est plus persistante que celle des tropiques ?
Les perturbations impliquent aussi un changement dans l’abondance des espèces où elles peuvent entraîner une suppression de certaines d’entre elles et une modification des conditions environnementales. Suivant leurs intensités l’effet sur l’écosystème sera visible à plus ou moins le long terme. Certains systèmes sont peu modifiés par les perturbations et donc sont plus résistants, persistants et moins variables. Dans les tropiques, les populations ne varient pas moins que les écosystèmes tempérés mais varient plus dans les systèmes climatiquement imprévisibles que les prévisibles, ce qui suggère que ces facteurs externes régissent la variabilité et influencent le système. Des modèles testent l’effet de la suppression d’espèce et la complexité favorise la stabilité et l’effet de la perte d’une espèce sera atténué. Sur de courte période les compositions en espèces sont constantes et les systèmes complexes sont favorisés face à la perte d’espèce, mais sur le long terme un changement des conditions environnementales suite à cette perte sera observé. Il est donc très important de prendre en compte l’échelle temporelle et spatiale dans l’étude de la relation complexité stabilité d’un écosystème.
Si un écosystème est plus stable lorsqu'il est complexe il semblerait évident que son évolution conduirait à sa complexification augmentant ainsi sa diversité spécifique et ses interactions. De plus des écosystèmes présentant peu d'espèces sont susceptibles d'être colonisés facilement par de nouvelles espèces et donc de se complexifier sur le court terme. Les échelles temporelles et spatiales sont importantes à prendre en compte, les écosystèmes sont soumis à des pressions environnementales, engendrant à court terme au mieux une modification des interactions et de l'abondance en espèce et au pire une disparition de certaines d'entre elles. Il semblerait qu'avec des conditions optimales à l'instar des régions tropicales les écosystèmes se complexifieraient. Cependant les perturbations pourraient impacter négativement ces écosystèmes. Ces perturbations contrôlent elles l'évolution des écosystèmes et limitent elles leur complexification ?
Des écosystèmes sont peu modifiés, ceci implique t il un "arrêt" de l'évolution à partir d'un certain degré de complexification ?
La présence d'écosystèmes complexes et simples sous entend elle que les systèmes simple sont une "transition" vers des systèmes plus complexe ?
Un écosystème déjà complexe va t il continuer à se complexifier ?
Il a été montré que des écosystèmes diversifiés étaient moins sujets à la colonisation par d'autres espèces, cela sous entend il qu'il existerait une limite a la complexification ?
Des systèmes trophiques complexes sont plus fragiles et sujets aux extinctions que les systèmes simples et donc ont tendance à durer sur une courte période, cela peut sous entendre que l'on observerait une complexification du système sur le court terme mais peut être une simplification sur le long terme suite à la fragilité du système
Dernière modification il y a plus de 10 ans.
De l'extracellulaire à l'intracellulaire : L'établissement des mitochondries et des choloplastes
From Extracellular to Intracellular: The Establishment of Mitochondria and Chloroplasts
J. M. Whatley, P. John, F. R. Whatley
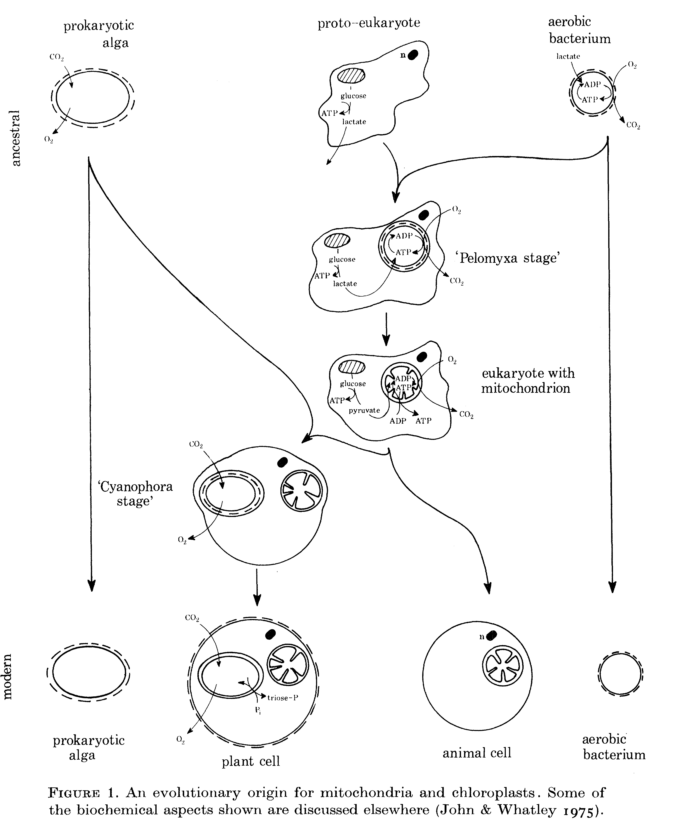
Origine évolutive des mitochondries et des chloroplastes (John & Whatley, 1975)
La théorie des endosymbioses multiples propose qu'un eucaryote primitif ait successivement phagocyté une bactérie et une cyanobactérie menant respectivement vers l'obtention de mitochondries et de chloroplastes. Cette évolution des procaryotes vers les eucaryotes semble donc s'être accompagnée d'une certaine complexification des organites cellulaires.
Les théories concernant l'évolution à l'échelle cellulaire sont particulièrement difficiles à tester en raison du manque de preuves fossiles. Dans cette publication (assez ancienne) ils se sont donc concentrés sur des preuves basées sur des organismes actuels. Certaines bactéries aérobies peuvent être considérées comme potentiels ancêtres des mitochondries, certaines algues procaryotes sont raisonablement considérées comme ancêtres des chloroplastes et _Pelomyxa palustris _serait possiblement l'hôte ancestral.
Cet article est une étude comparative des organismes cellulaires connus et de leurs organites. Il se penche sur
- la comparaison de Paracoccus denitrificans et des mitochondries ( comparaison de la chaine de réaction, du cytochrome C, du transfert de gènes
- l'étude de l'hydrogénosome des Trichomonas anaérobies
- la comparaison des bactérie endosymbiotiques et des mitochondries (modèle de l'amibe géante Pelomyxa palustris)
- l'étude des procaryotes photosynthétiques et de leurs chloroplastes (pigments et structure thylacoïde)
- l'évolution des chloroplastes chez les Rhodophytes, Chlorophytres, plantes terrestres, et Chromophytes
La théorie des endosymbioses en série propose traditionnellement que la mitochondrie et le chloroplaste aient évolués à partir de procaryotes unicellulaires libres. Cette étude comparative a mis en évidence et résumé certaines preuves biochimiques et structurelles en faveur de cette théorie. En particulier, il a été discuté du fait que Prochloron et Paracoccus puissent être des représentations modernes de l'ancêtre supposé des chloroplastes et des mitochondries, tandis que Cyanophora et Pelomyxa peuvent être considérées comme de possibles formes intermédiaires entre cet ancêtre et les organites actuels.
Il a ensuite été proposé que cette théorie soit modifiée en ce qui concerne les chloroplastes de Dinophycées et d'Euglénophycées, qui ont probablement évolués à partir de symbiontes chololastiques isolées, et que les unités chloroplastiques d'autres algues chromophytes aient évolués de symbiontes eucaryotes (figure 15).
La théorie des endosymbioses multiples permet d'expliquer la transition procaryote/eucaryote et est très supportée par les connaissances actuelles des organismes cellulaires, leurs organites, leur structure, leur fonctionnement et leur composition biochimique.
Ainsi, on peut considérer que l'évolution des eucaryotes correspond bien à une complexification par rapport aux procaryotes en raison de l'apparition de nouveaux organites par endosymbioses successives, permettant l'acquisition de nouvelles fonctions.
Néanmoins, cet article est assez ancien et les techniques modernes d'analyse de séquences ADN, d'inférence phylogénétique, de modélisation informatique etc. permettent aujourd'hui de renforcer ou d'invalider certains aspects de cette théorie et de construire les arbres phylogénétiques les plus probables.[1]
Dernière modification il y a plus de 10 ans.
L'évolution implique-t-elle la complexification ?
En biologie, l'évolution peut être définie comme la différence morphologique et génétique observée d’une génération à l’autre. A un sens plus large, elle englobe non seulement les organismes, les populations, les espèces mais également les écosystèmes (successions écologiques par exemple).
Publiée il y a environ 6 ans par Un illustre inconnu.Elle consiste donc au changement de caractère au fil des générations. Les caractères peuvent être considérés à différentes échelles : moléculaires, cellulaires, physiologiques, individuelles, populationnelles et écosystémiques.
Il est important de prendre en compte ces différentes échelles lorsque l’on s'interroge sur la finalité de l’évolution au sens large comme au sens strict.
Il est souvent considéré que l'évolution est gage de complexification. Cette dernière peut être considéré comme l'élaboration progressive de solutions par l'ajout de nouvelle structure.
L'évolution aboutit-elle obligatoirement à une complexification du système, observe-t-on au contraire, une simplification au cours des génération ? Y a-t-il une tendance globale ou bien le nombre de réponse à cette question dépend du nombre de types d’organismes vivants ?
Dernière modification il y a environ 6 ans.