Vous venez de mettre les pieds dans les tréfonds en construction de ControverSciences.
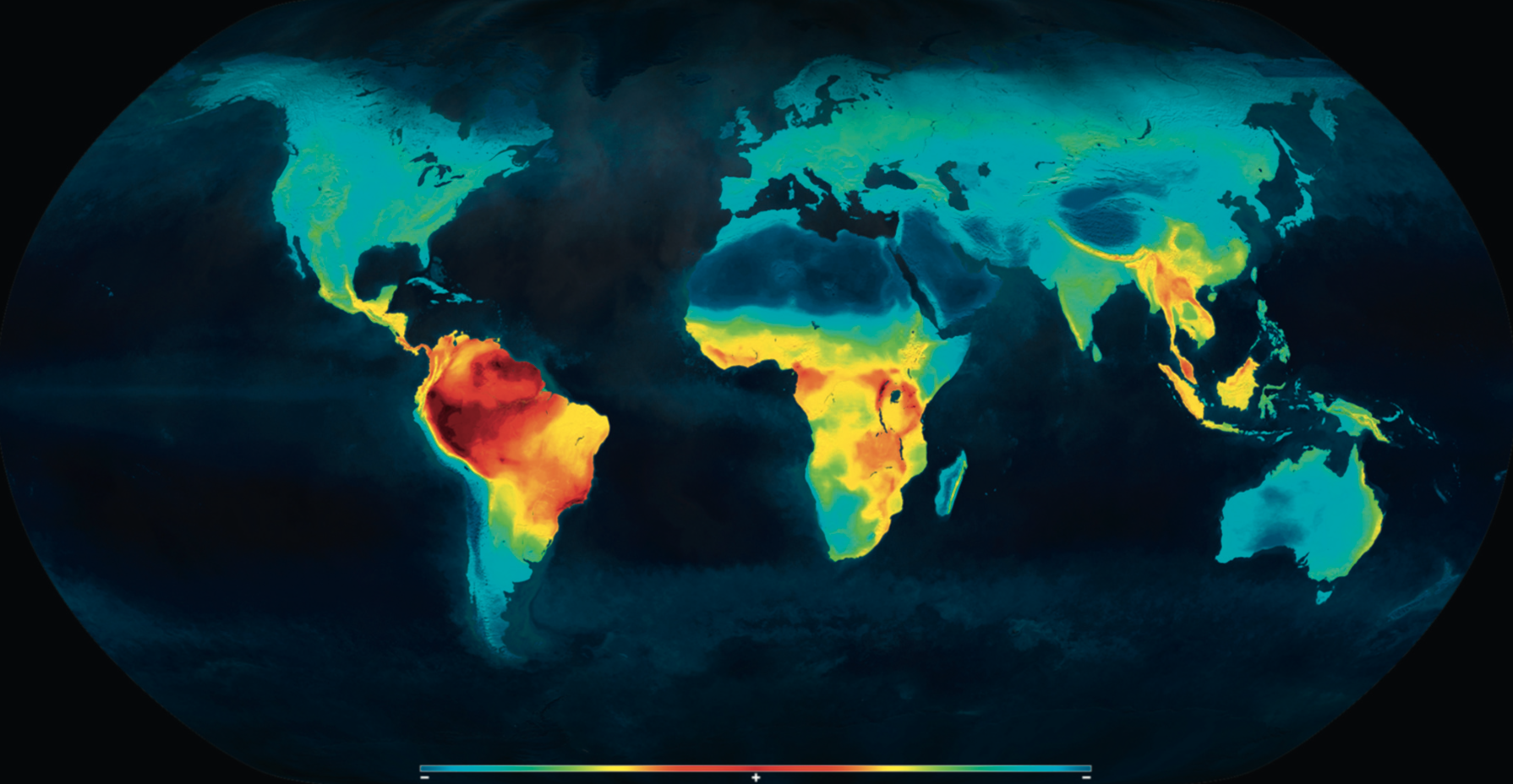
Planisphère montrant la distribution des vertébrés terrestres actuels. Une diversité plus importante s'observe ainsi au niveau des zones équatoriales.
Introduction
Le gradient latitudinal de biodiversité (GLB) est un phénomène macroécologique majeur en biogéographie. En effet, Wallace en fait la première description en 1876, dans son ouvrage « The Geographical Distribution of Animals ». Depuis lors, l’hétérogénéité des patrons de distribution spatiale de la biodiversité n’a eu cesse d’alimenter les débats scientifiques.
Malgré l’acceptation du caractère ubiquiste de ce phénomène de décroissance des valeurs de la biodiversité en fonction de la latitude, de nombreuses hypothèses ont été émises afin de rendre compte de la formation d’un tel gradient (Hawkins et al, 2003; Hillebrand, 2004; Valentine et al, 2008; Wright et al, 2006). De plus, des contre-exemples à la diminution de la biodiversité en fonction de la latitude sont référencés (Mateo et al, 2016; Visser et al, 2013; Boyero et al, 2011; Rolland et al, 2014). A cela s’ajoutent des données paléontologiques contrastant l’universalité du GLB dans le temps long (Mannion et al, 2014; Mannion et al, 2015; Rolland et al, 2015; Marcot et al, 2016).
Devant ces contradictions, nous proposons une revue approfondie des débats en cours concernant le gradient latitudinal de biodiversité. Nous nous attacherons ainsi à décrire les hypothèses postulées quant à la formation du GLB, à exposer les contre-exemples au GLB et enfin à expliciter les méthodologies employées dans l’étude d’un tel gradient.
Coeur de la synthèse
Le déclin de la biodiversité des basses latitudes vers les hautes latitudes est un phénomène applicable à une large gamme d’organismes vivants (Hawkins et al, 2003; Condamine et al, 2012; Böhm et al, 2005; Andam et al, 2016; Siqueira et al, 2016; Rolland et al, 2014; Marcot et al, 2016; Mateo et al, 2016).
Les hypothèses responsables de cette variation latitudinale de biodiversité sont néanmoins débattues (Hillebrand, 2005). Ces hypothèses font appel à des processus écologiques et évolutifs variés (Visser et al, 2013; Condamine et al, 2012; Rolland et al, 2014) mais qui dépendent tous d’une cause proximale en lien avec des facteurs climatiques, historiques et géographiques (Mannion et al, 2014). La variabilité de l’intensité d’insolation et l’impact de la saisonnalité sont des facteurs climatiques prépondérants (Mannion et al, 2015; Condamine et al, 2012; Visser et al, 2013) qui contrastent avec les caractéristiques géographiques et topographiques des régions considérées (Böhm et al, 2005; Kneitel et al, 2015). À ces paramètres s’ajoute une dimension historique rendant compte de l’effet des perturbations et des événements abiotiques passés sur la biodiversité actuelle (Böhm et al, 2005; Mannion et al, 2014; Mannion et al, 2015).
Ces facteurs vont ainsi impacter de nombreux phénomènes écologiques et évolutifs, eux-mêmes responsables des différentiels de distribution spatiale de la biodiversité. On peut ainsi mettre en lumière les phénomènes de conservation des niches écologiques (Fernandez et al, 2016), de dispersion et de migration (Böhm et al, 2005; Rolland et al, 2015), d’exclusion compétitive et d’agrégation des niches écologiques (Siqueira et al, 2016), de diversification (taux d’extinction et de spéciation) (Visser et al, 2013; Rolland et al, 2014; Andam et al, 2016) ou encore d’augmentation de taux d’évolution moléculaire (Wright et al, 2006) dans la formation d’un gradient latitudinal de diversité.
Même si la primauté du facteur climatique est souvent mise en avant (Mannion et al, 2015), la mise en place d’un gradient de diversité n’est pas à chercher du côté d’un seul facteur mais plutôt au sein d’une synergie entre les différents facteurs mentionnés précédemment (Mannion et al, 2014; Marcot et al, 2016; Condamine et al, 2012).
Toutefois, certains taxa semblent faire exceptions à la règle du gradient latitudinal de diversité (Hillebrand, 2004). On notera ainsi des diversités plus importantes aux latitudes tempérées pour les invertébrés détritivores (Boyero et al, 2011), les bryophytes (Mateo et al, 2016), certains clades d’oiseaux (Hillebrand, 2004), un ordre de mammifères (Rolland et al, 2014) et enfin certains taxons de graminées (Visser et al, 2013) . De nombreuses hypothèses écologiques et évolutives en référence aux facteurs climatiques, géographiques et historiques ont été avancées afin de rendre compte de ces exceptions.
Cependant, les critiques les plus virulentes concernant l’universalité du gradient latitudinal de diversité ne viennent pas des données néontologiques mais des données paléontologiques. En effet, des études approfondies des données d’occurrences spatiales et temporelles de divers taxa fossiles ont permis de démontrer le caractère ponctuel du GLD dans le temps long (Mannion et al, 2014; Mannion et al, 2015; Marcot et al, 2016; Rolland et al, 2015). La formation d’un tel gradient semble ainsi faire écho à des périodes de chute globale des temperatures, mettant ainsi en exergue l’importance du facteur climatique. Cependant, le rôle des facteurs géographiques et historiques n’est pas à négliger (Böhm et al, 2005; Condamine et al, 2012). Enfin, il est important de mettre en lumière les différences de comportement des faunes marines et terrestres vis à vis d’une distribution latitudinale de la biodiversité (Mannion, 2014; Mannion, 2015).
Afin de contraster ces différents points de vues, il nous parait essentiel de s’intéresser aux différentes méthodologies employées dans l’étude du GLB. La diversité peut ainsi être appréhendée de différentes manières. La diversité taxinomique se trouve être au coeur de nombreuses études (Fernandez et al, 2016; Kneitel et al, 2015; Visser et al, 2013) tandis que l’étude de la diversité phylogénétique semble se démocratiser (Condamine et al, 2012; Andam et al, 2016; Siqueira et al, 2016; Rolland et al, 2014; Rolland et al, 2015). Enfin, les diversités écomorphologique (Lamanna et al, 2014) et génétique (Wright et al, 2006) complètent l’ensemble. Ces différentes méthodes apportent des résultats complémentaires mais parfois contradictoires concernant le gradient latitudinal de biodiversité. En effet, les études portant sur les adaptations morphofonctionnelles (Lamanna et al, 2014) et écomorphologiques (Boyero et al, 2011) mettent en contraste une diversité taxinomique forte aux tropiques mais une variabilité de stratégies adaptatives plus importante au niveau des latitudes tempérées. Enfin les analyses de diversité phylogénétique permettent de mettre en lumière des événements de diversification au sein de clades des latitudes tempérées et tropicales. Les corrélations existantes entre ces événements de diversifications et des paramètres biotiques ou abiotiques peuvent nous renseigner sur l’importance des facteurs climatique, géographique et historique dans la mise en place du GLB (Condamine et al, 2012; Böhm et al, 2005).
La limite de chacune des méthodes considérées est donc un paramètre primordial dans l’appréhension de la validité et de l’universalité du gradient latitudinal de diversité.
Conclusion et ouverture
La remise en question du gradient latitudinal de diversité est au coeur de cette synthèse. L’aspect multidisciplinaire de l’étude du GLB démontre bien les multiples facettes présentées par ce phénomène. En effet, l’existence actuelle d’un gradient latitudinal de diversité ne fait aucun doute tandis que l’universalité d’un tel gradient dans le temps long semble beaucoup plus contestable. A cela s’ajoute un panel conséquent d’hypothèses évolutives et écologiques, renforçant ainsi les incertitudes liées aux origines de la mise en place d’un tel patron de distribution de la biodiversité.
La compréhension du comportement des faunes passées et actuelles devant les fluctuations des paramètres biotiques et abiotiques considérées dans cette synthèse paraît primordiale à la lumière des changements climatiques qui s’annoncent. Les études biogéographiques prennent ainsi une place de choix dans la compréhension des faunes à la fois contemporaines et anciennes.
Dernière modification il y a plus de 6 ans.
Cette synthèse se base sur 19 références.
- • L'énergie, l'eau et les profils géographiques à grande échelle de la richesse spécifique
- • Généralités sur le gradient latitudinal de biodiversité.
- • Facteurs biotiques, diversité et gradient de latitude.
- • Un tempo évolutif plus rapide au niveau des zones tropicales
- • Les mousses "nordiques" : un gradient latitudinal de biodiversité inverse chez les bryophytes européens.
- • Mécanismes conduisant à un gradient latitudinal de diversité inhabituel pour les graminées
- • La distribution globale d'une guilde trophique majeure diffère du modèle de gradient latitudinal de diversité.
- • Une plus grande spéciation et une réduction de l'extinction dans les tropiques contribuent à la mise en place du gradient latitudinal de diversité des mammifères
- • Le gradient latitudinal de biodiversité à travers les âges
- • Le climat contraint l'histoire évolutive et la biodiversité des crocodiliens
- • La dispersion est un des facteurs de la mise en place du gradient latitudinal de diversité chez les Carnivores
- • Mise en place tardive d'un gradient latitudinal de diversité au sein des mammifères nord-américains
- • Quelles sont les causes du gradient latitudinal de biodiversité ? Facteurs évolutifs et contraintes écologiques impliqués dans le processus de diversité des swallowtail.
- • Biogéographie historique et évolution du gradient latitudinal de diversité au sein des Papionini
- • Les bactéries du genre Streptomyces sont réparties en suivant un gradient latitudinal de diversité.
- • Processus évolutifs sous-jacents dans les différentes latitudes chez les poissons de récif.
- • Les facteurs climatiques modifient la taille des habitats et déterminent le gradient de diversité dans les mares temporaires.
- • Etude du gradient des amibes testacées dans le sud-ouest de l'Amérique du sud : équilibre eau-énergie et évolution.
- • Diversité fonctionnelle et gradient latitudinal de diversité
Les mousses "nordiques" : un gradient latitudinal de biodiversité inverse chez les bryophytes européens.
The mossy north: an inverse latitudinal diversity gradient in European bryophytes
Rubén G. Mateo, Olivier Broennimann, Signe Normand, Blaise Petitpierre, Miguel B. Araújo, Jens-C. Svenning, Andrés Baselga, Federico Fernández-González, Virgilio Gómez-Rubio, Jesús Muñoz, Guillermo M. Suarez, Miska Luoto, Antoine Guisan, Alain Vanderpoorten
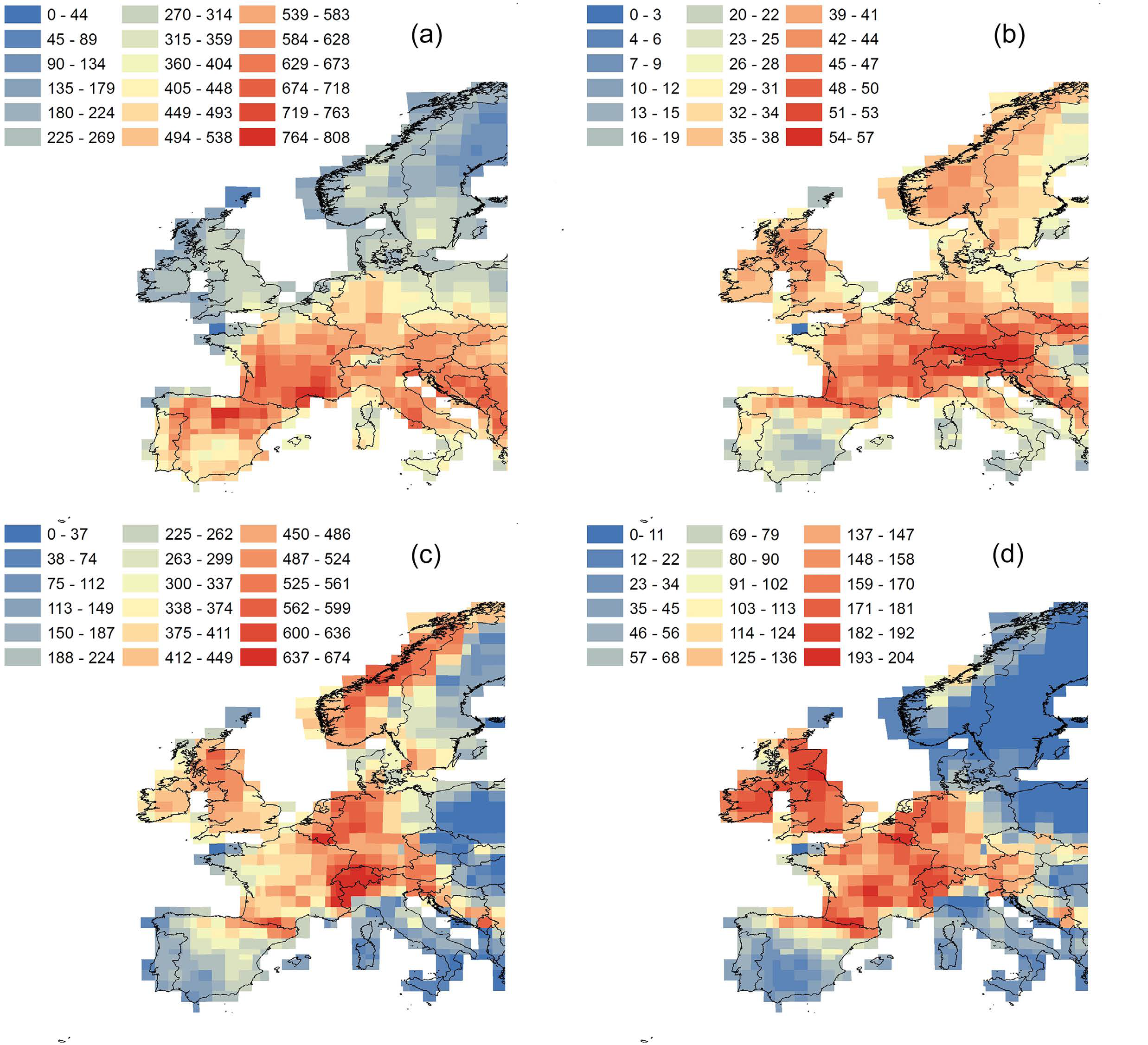
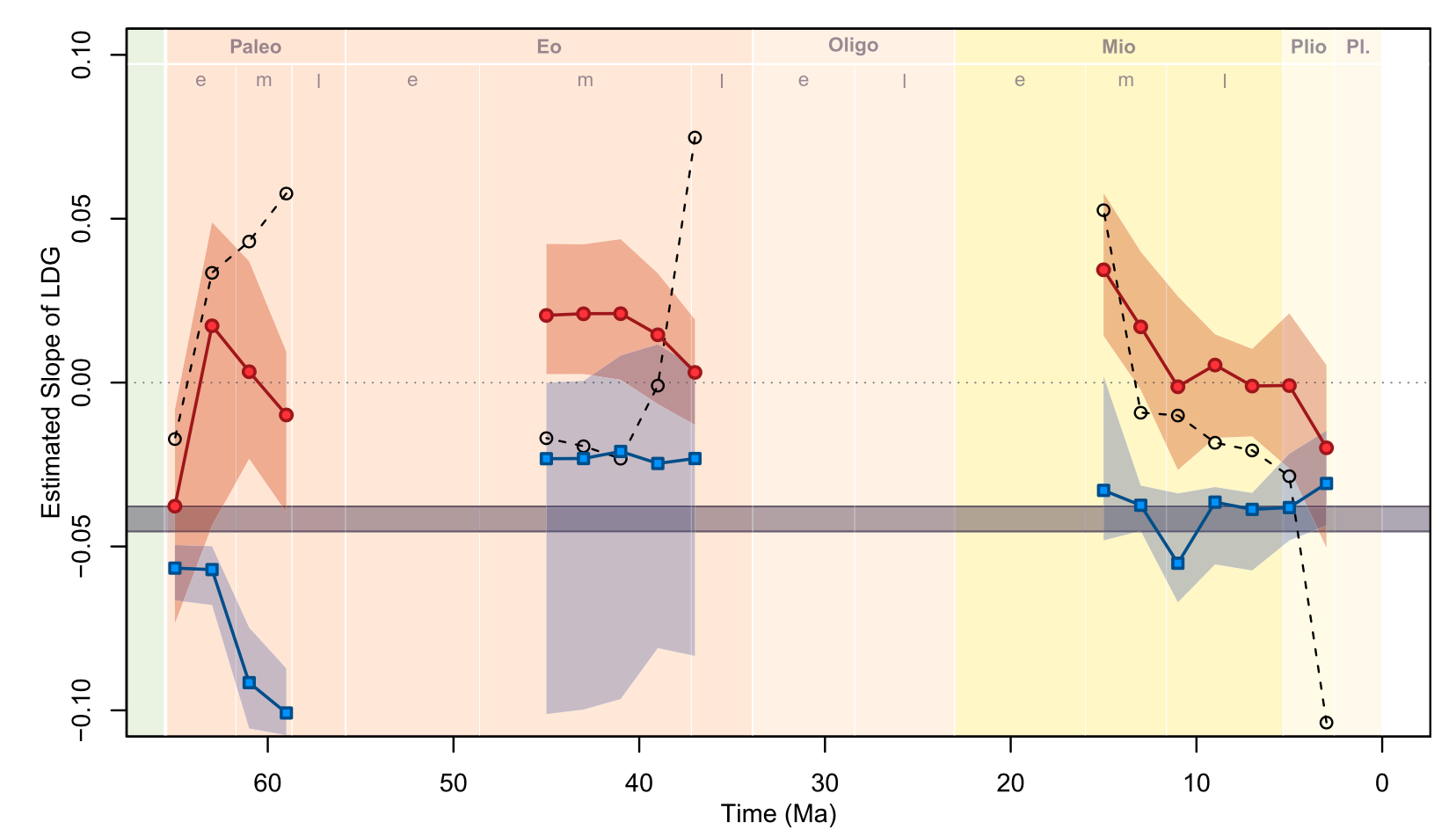
Mise en place tardive d'un gradient latitudinal de diversité au sein des mammifères nord-américains
Late Cenozoic onset of the latitudinal diversity gradient of North American mammals
Jonathan D.Marcot , David L.Fox, Spencer R.Niebhur
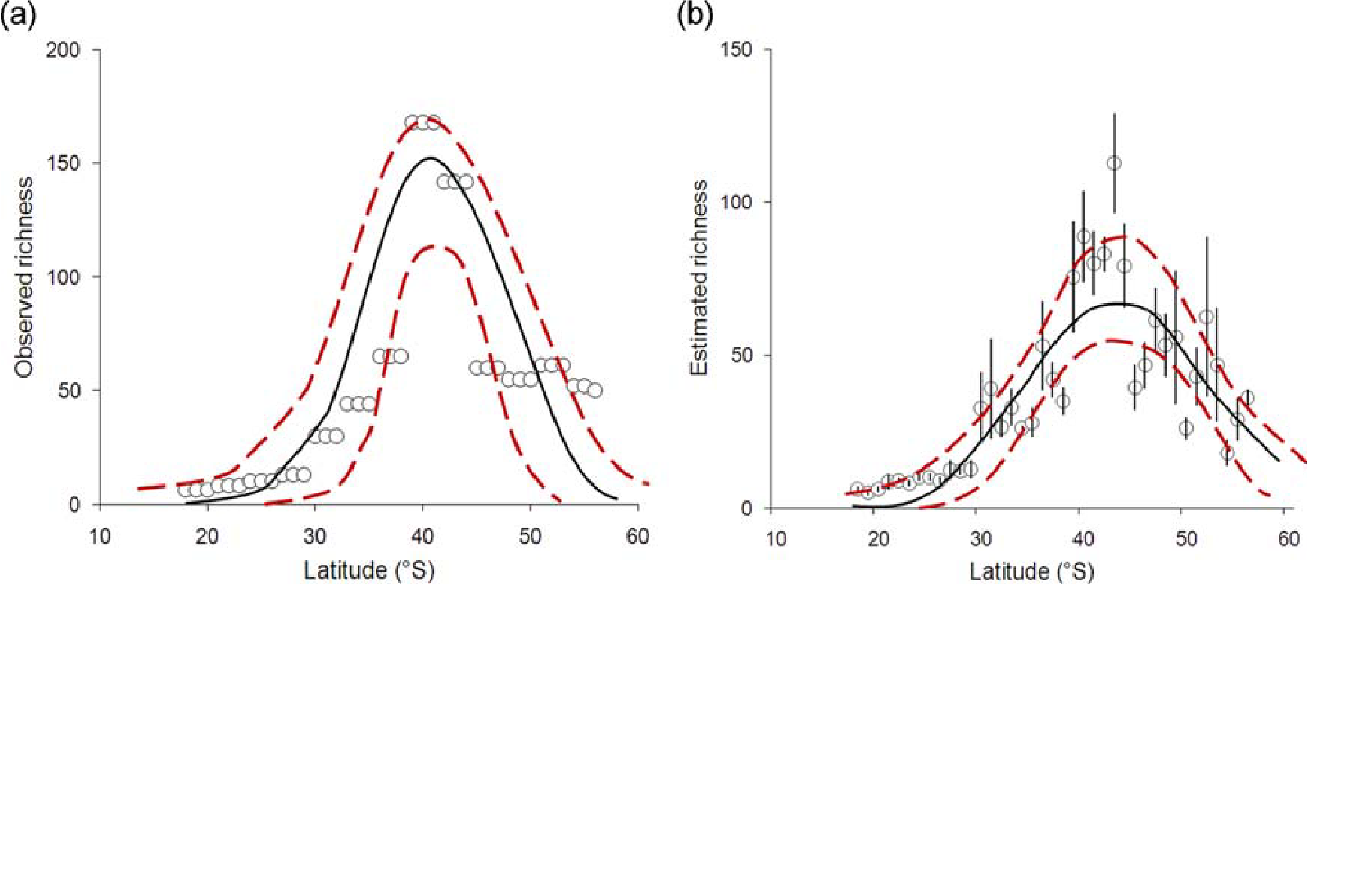
Etude du gradient des amibes testacées dans le sud-ouest de l'Amérique du sud : équilibre eau-énergie et évolution.
Water–energy balance, past ecological perturbations and evolutionary constraints shape the latitudinal diversity gradient of soil testate amoebae in south-western South America
Leonardo D. Fernandez, Bertrand Fournier, Reinaldo Rivera, Enrique Lara, Edward A. D. Mitchell, and Cristian E. Hernandez
Processus évolutifs sous-jacents dans les différentes latitudes chez les poissons de récif.
Evolutionary processes underlying latitudinal differences in reef fish biodiversity
Alexandre C. Siqueira, Luiz Gustavo R. Oliveira-Santos, Peter F. Cowman, Sergio R. Floeter
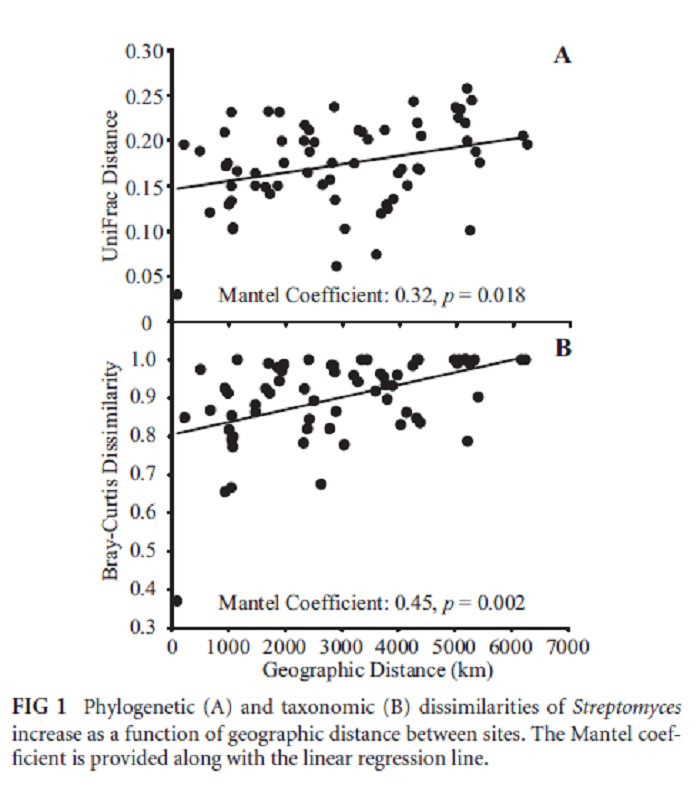
Les bactéries du genre Streptomyces sont réparties en suivant un gradient latitudinal de diversité.
A Latitudinal Diversity Gradient in Terrestrial Bacteria of the Genus Streptomyces
Cheryl P. Andam, James R. Doroghazi, Ashley N. Campbell, Peter J. Kelly, Mallory J. Choudoir, Daniel H. Buckley
Les facteurs climatiques modifient la taille des habitats et déterminent le gradient de diversité dans les mares temporaires.
Climate-driven habitat size determines the latitudinal diversity gradient in temporary ponds
Jamie M. Kneitel
La dispersion est un des facteurs de la mise en place du gradient latitudinal de diversité chez les Carnivores
Dispersal is a major driver of the latitudinal diversity gradient of Carnivora
Jonathan Rolland, Fabien L. Condamine, Champak R. Beeravolu, Frédéric Jiguet, Hélène Morlon
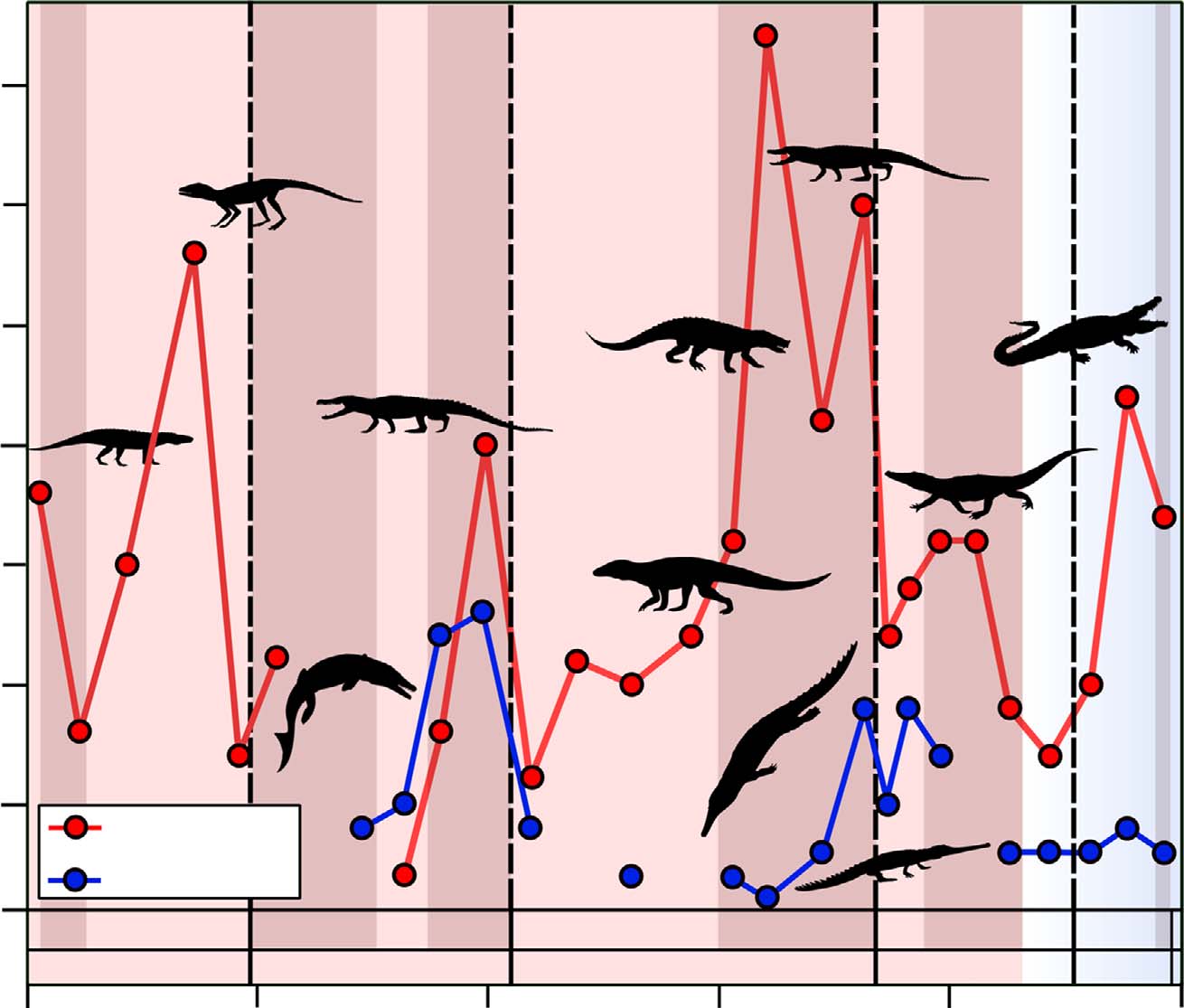
Le climat contraint l'histoire évolutive et la biodiversité des crocodiliens
Climate constrains the evolutionary history and biodiversity of crocodylians
Philip D. Mannion, Roger B. J. Benson, Matthew T. Carrano, Jonathan P. Tennant, Jack Judd, Richard J. Butler
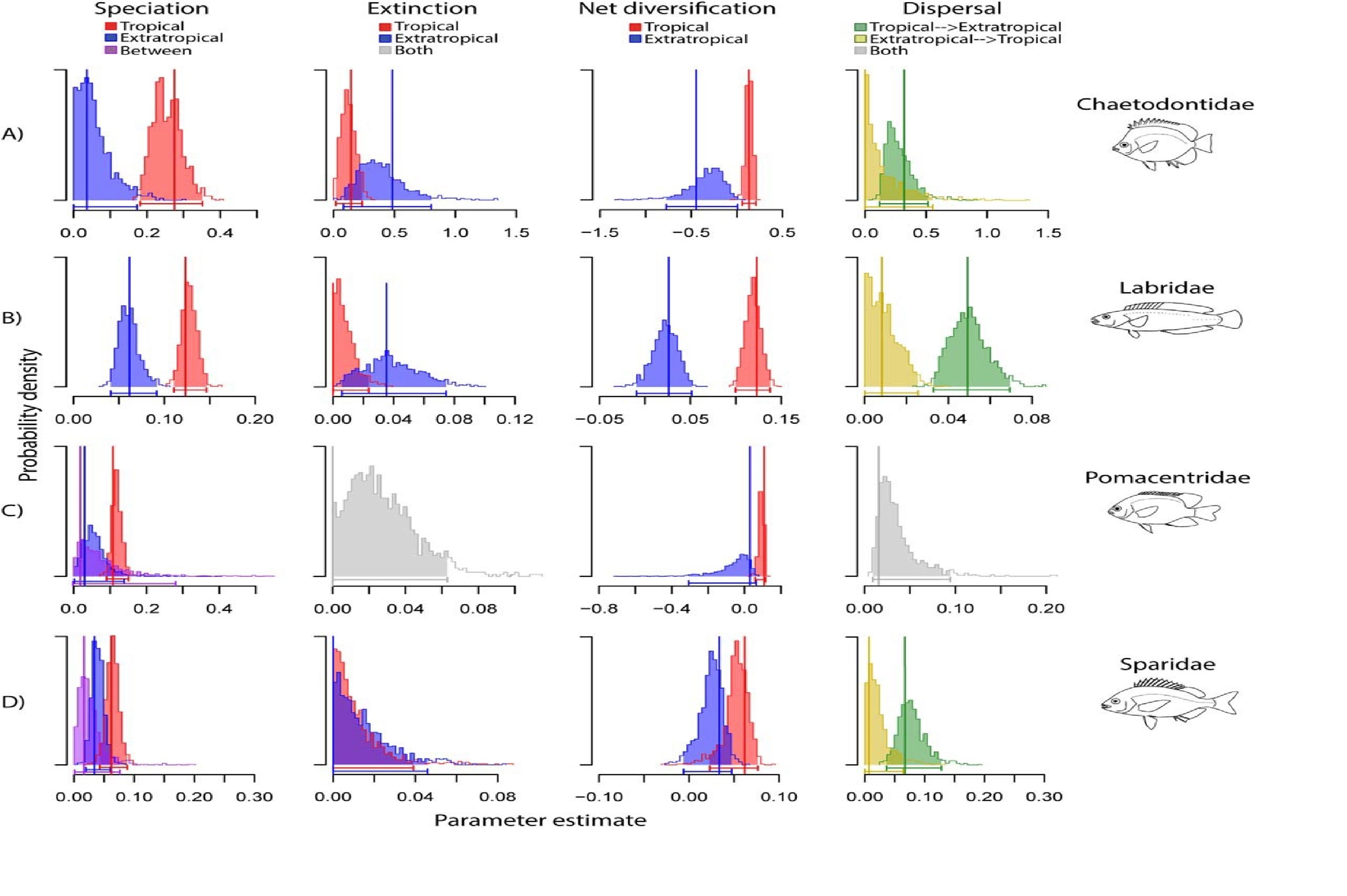
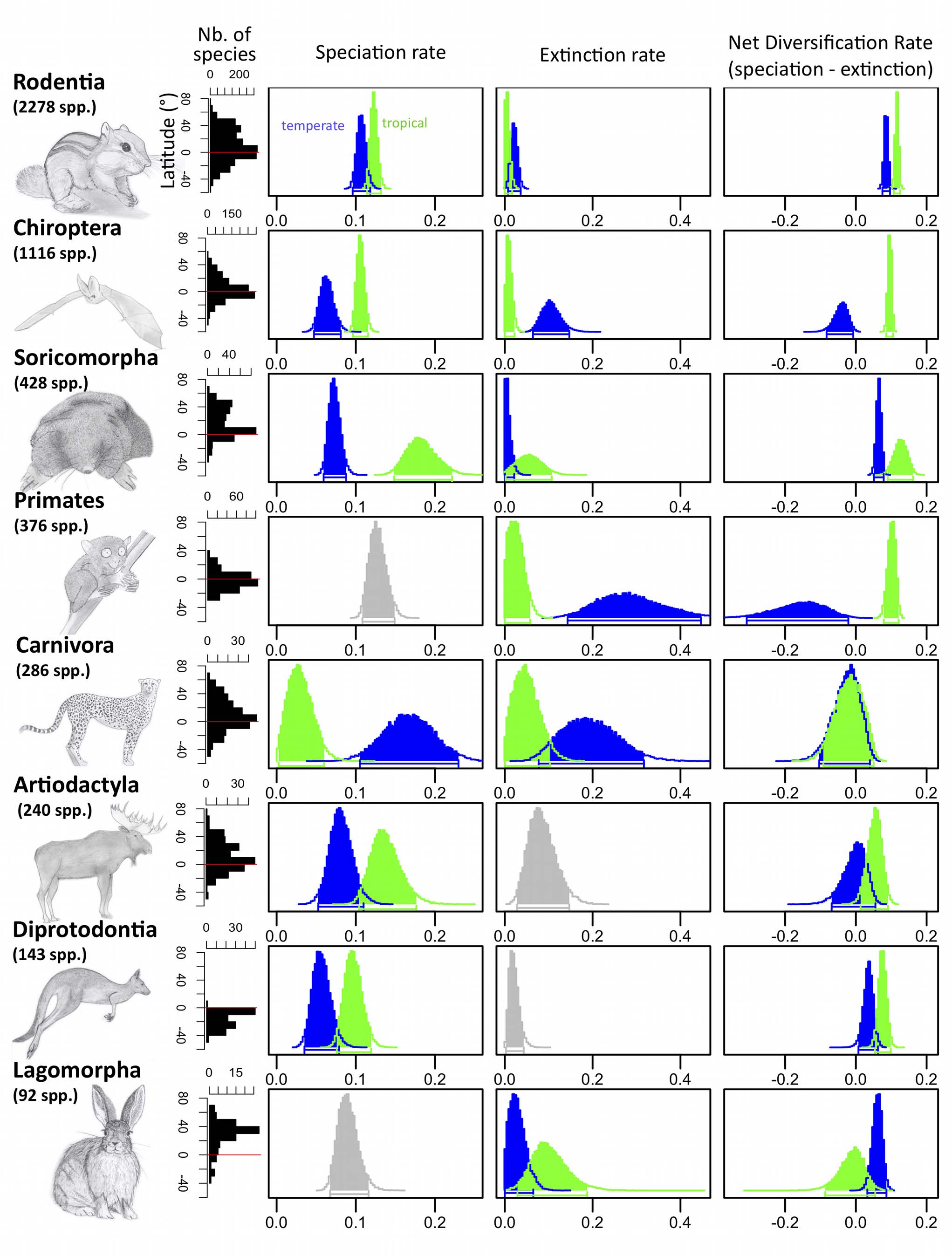
Une plus grande spéciation et une réduction de l'extinction dans les tropiques contribuent à la mise en place du gradient latitudinal de diversité des mammifères
Faster Speciation and Reduced Extinction in the Tropics Contribute to the Mammalian Latitudinal Diversity Gradient
Jonathan Rolland, Fabien L. Condamine, Frederic Jiguet, Hélène Morlon
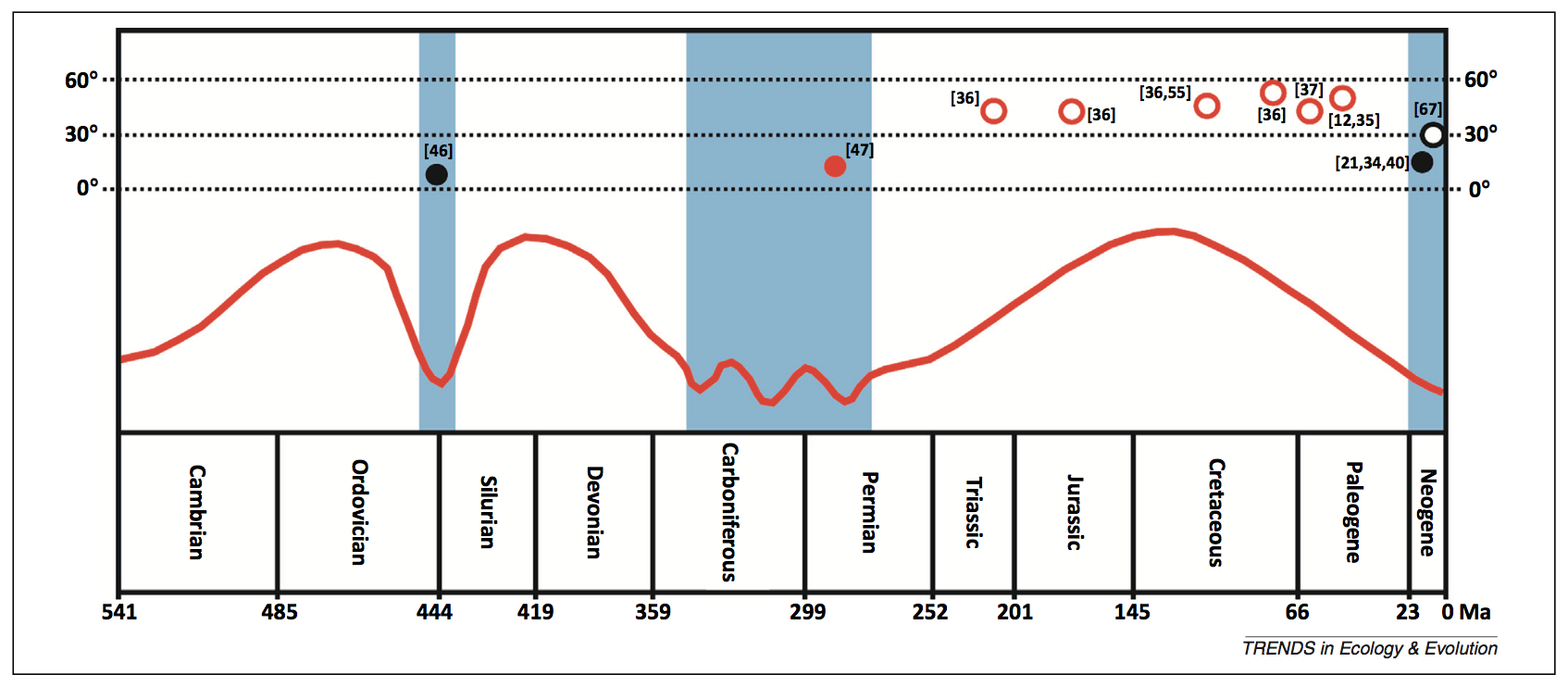
Le gradient latitudinal de biodiversité à travers les âges
The latitudinal biodiversity gradient through deep time
P.D. Mannion, P. Upchurch, R.B.J. Benson, A. Goswami
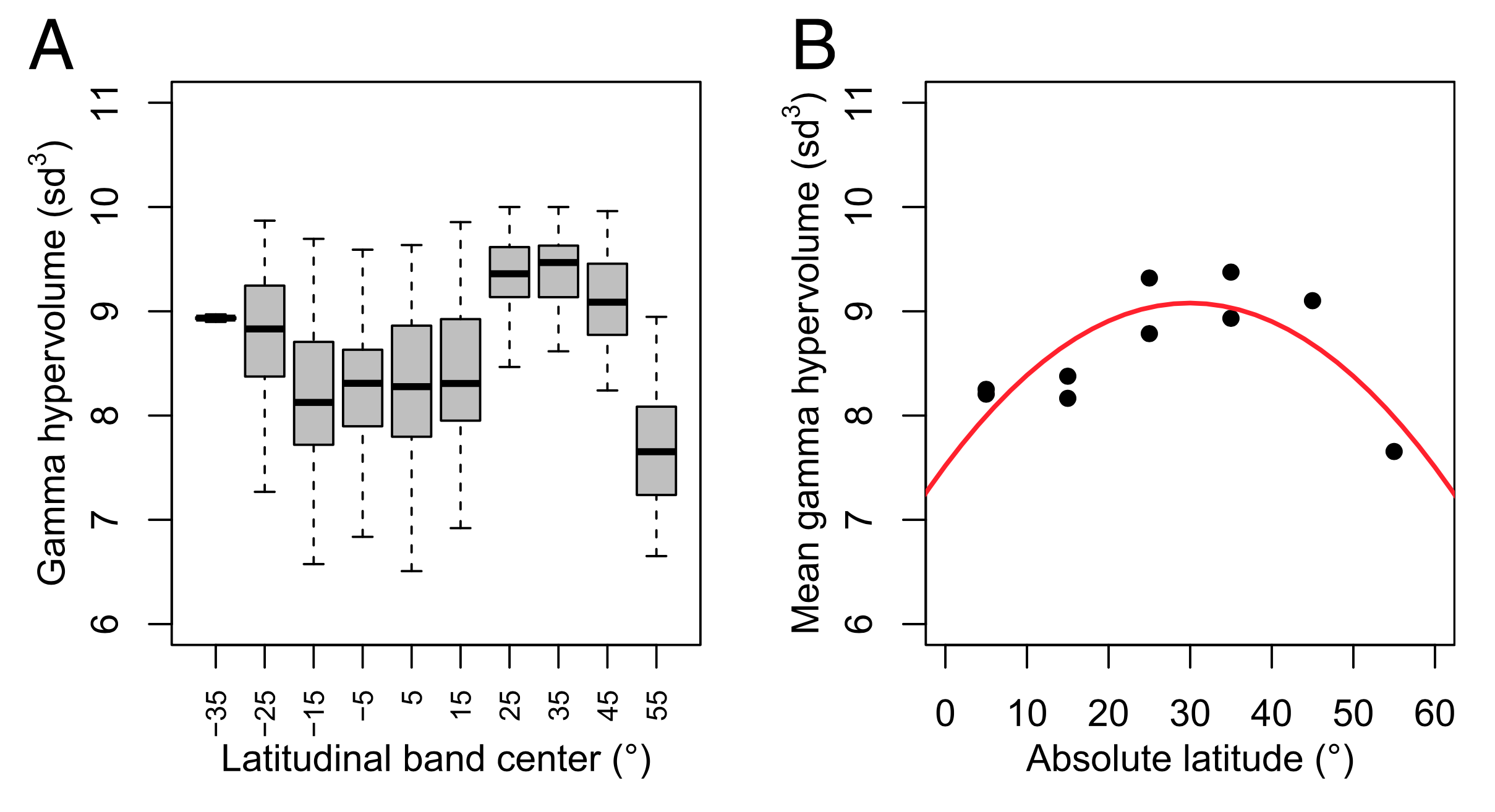
Diversité fonctionnelle et gradient latitudinal de diversité
Functional trait space and the latitudinal diversity gradient
C.Lamanna et al
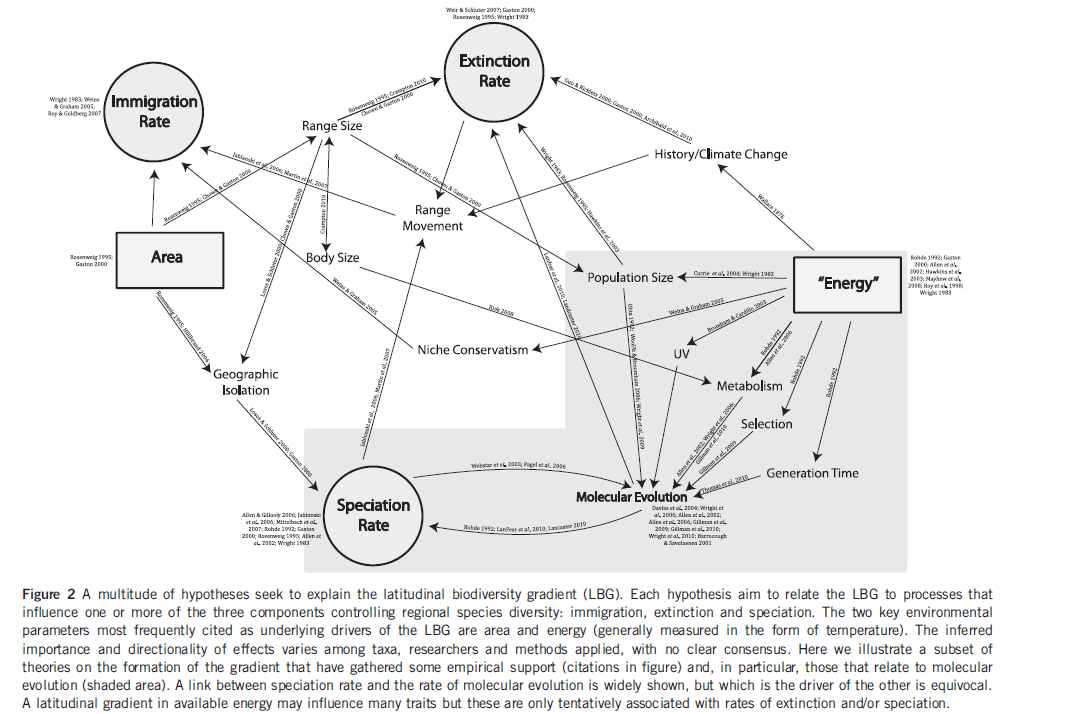
Evolution moléculaire et Gradient latitudinal de Biodiversité
Molecular evolution and the latitudinal biodiversity gradient
EJ Dowle, M Morgan-Richards and SA Trewick
Mécanismes conduisant à un gradient latitudinal de diversité inhabituel pour les graminées
Mechanisms driving an unusual latitudinal diversity gradient for grasses
Vernon Visser, W. Derek Clayton, David A. Simpson, Robert P. Freckleton, Colin P. Osborne
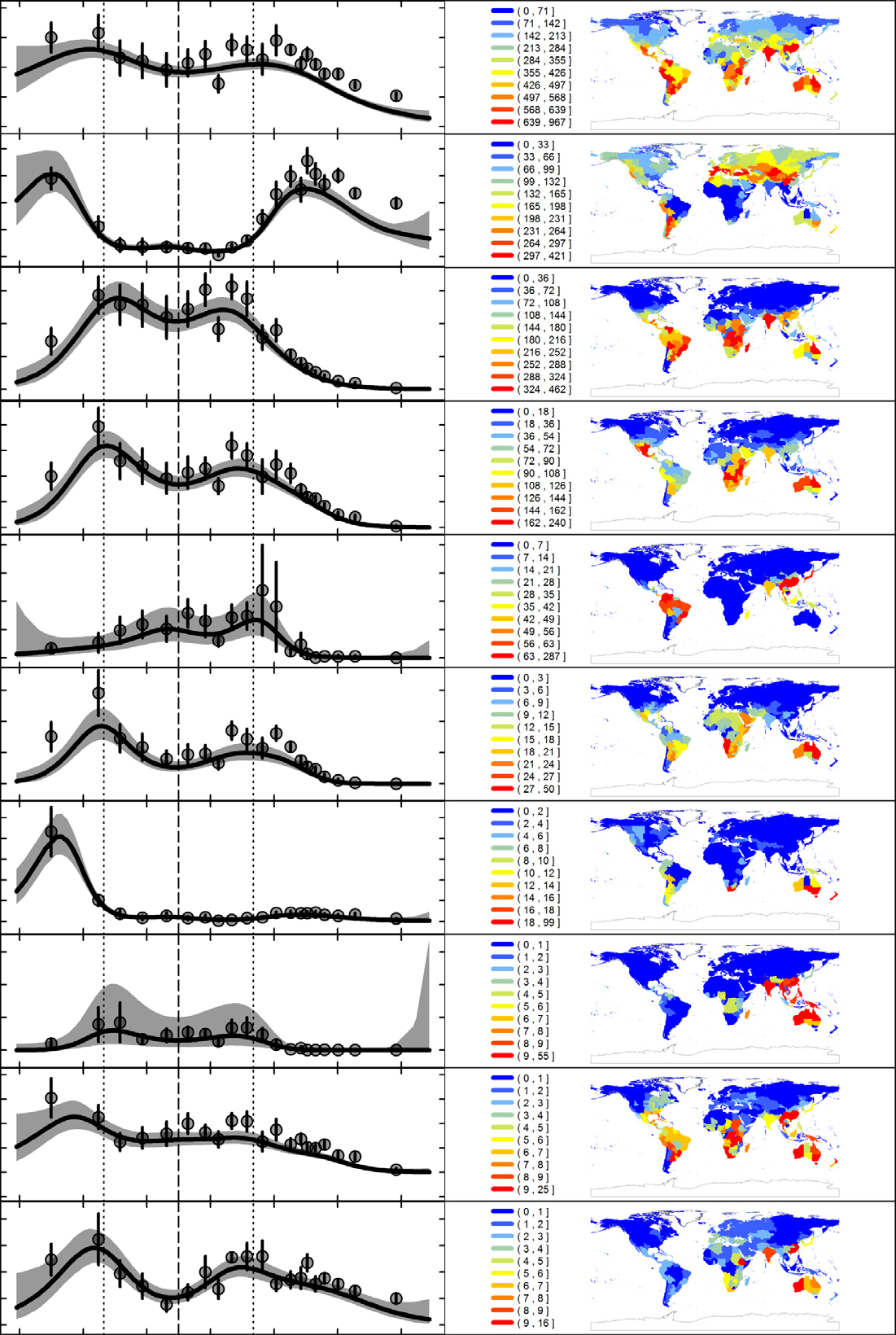
Quelles sont les causes du gradient latitudinal de biodiversité ? Facteurs évolutifs et contraintes écologiques impliqués dans le processus de diversité des swallowtail.
What causes latitudinal gradients in species diversity? Evolutionary processes and ecological constraints on swallowtail biodiversity
Fabien L. Condamine, Felix A. H. Sperling, Niklas Wahlberg, Jean-Yves Rasplus and Gael J. Kergoat
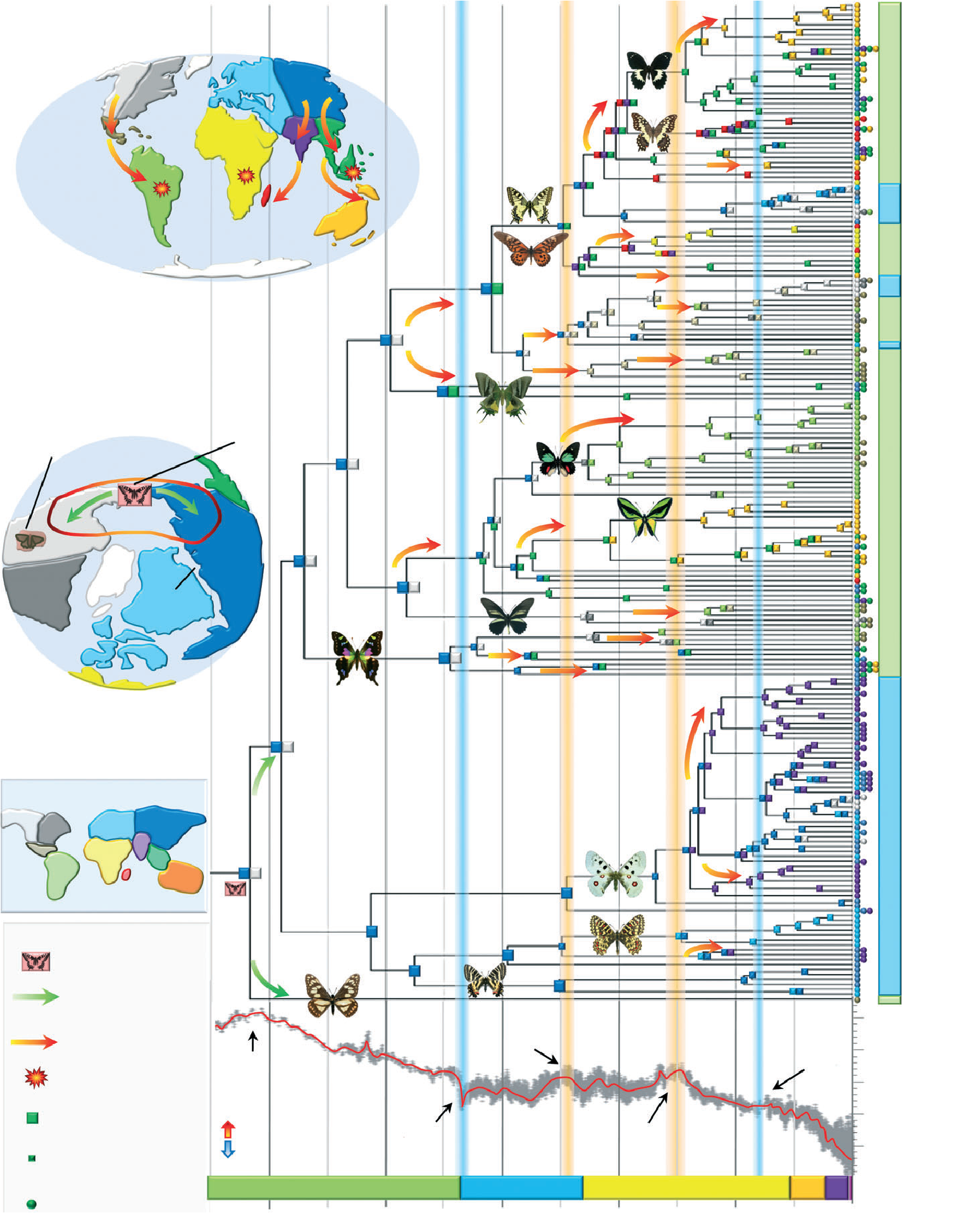
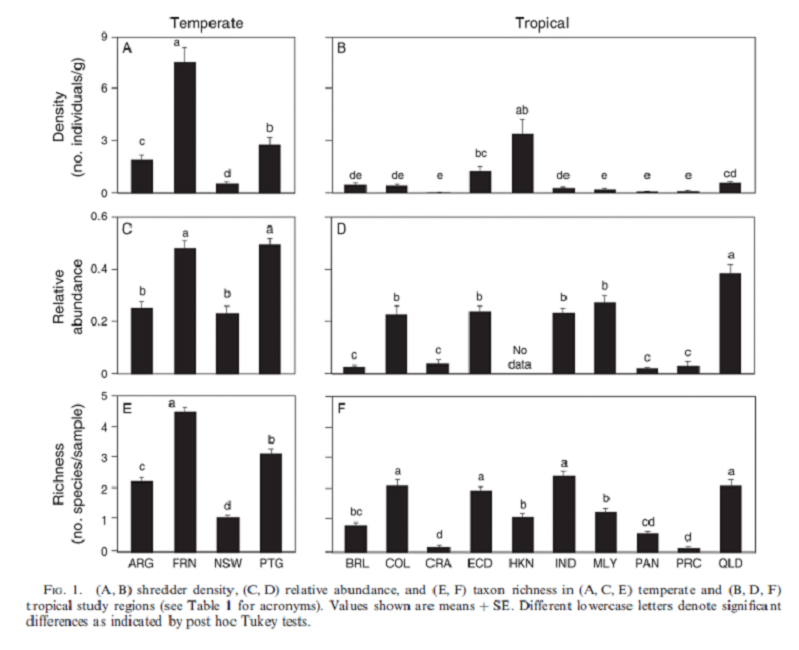
La distribution globale d'une guilde trophique majeure diffère du modèle de gradient latitudinal de diversité.
Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns
Luz Boyero, Richard G Pearson, David Dudgeon, Manuel A. S Graça, Mark O Gessner, Ricardo J Albariño, Verónica Ferreira, Catherine M Yule, Andrew J Boulton, Muthukumarasamy Arunachalam, Marcos Callisto, Eric Chauvet, Alonso Ramírez, Julián Chará, Marcelo S Moretti, José F Gonçalves, Julie E Helson, Ana M Chará-Serna, Andrea C Encalada, Judy N Davies, Sylvain Lamothe, Aydeè Cornejo, Aggie O. Y Li, Leonardo M Buria, Verónica D Villanueva, María C Zúñiga, Catherine M Pringle
Facteurs biotiques, diversité et gradient de latitude.
Incumbency, diversity, and latitudinal gradients
James W. Valentine, David Jablonski, Andrew Z. Krug, and Kaustuv Roy
Un tempo évolutif plus rapide au niveau des zones tropicales
The road from Santa Rosalia: A faster tempo of evolution in tropical climates
S. Wright, J. Keeling, L. Gillman
Biogéographie historique et évolution du gradient latitudinal de diversité au sein des Papionini
Historical biogeography and the evolution of the latitudinal gradient of species richness in the Papionini (Primata: Cercopithecidae)
M. Böhm, P.J. Mayhew
Généralités sur le gradient latitudinal de biodiversité.
On the Generality of the Latitudinal Diversity Gradient
Helmut Hillebrand
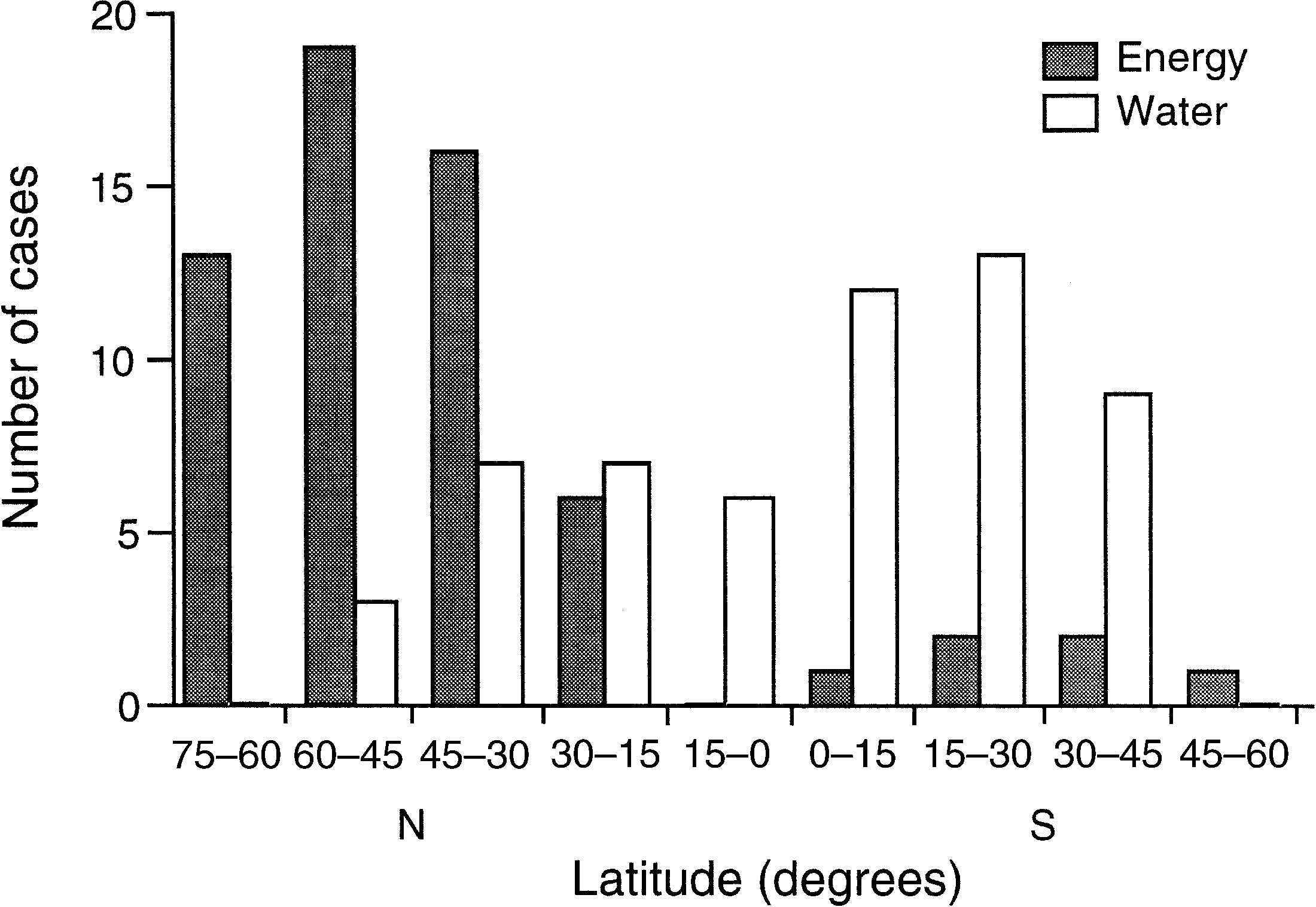
L'énergie, l'eau et les profils géographiques à grande échelle de la richesse spécifique
Energy, water, and broad-scale geographic patterns of species richness
Bradford A. Hawkins, Richard Field, Howard V. Cornell, David J. Currie, Jean-François Guégan, Dawn F. Kaufman, Jeremy T. Kerr, Gary G. Mittelbach, Thierry Oberdorff, Eileen M. O'Brien, Eric E. Porter and John R.G. Turner
Le gradient latitudinal de biodiversité est il universel et intemporel ?
L’observation d’une baisse de la biodiversité des zones équatoriales vers les zones septentrionales représente un phénomène macroécologique de premier ordre en biologie.
Ce gradient latitudinal de biodiversité fait consensus au sein de la communauté scientifique. Cependant, les causes de la mise en place d’un tel patron de distribution de la biodiversité restent encore largement débattues. De nombreuses hypothèses, alliant à la fois données paléontologiques et néontologiques, données écologiques et phylogénétiques, ont été mises en avant afin de rendre compte de la mise en place d’un gradient spatial de diversité.
Malgré cela, de nombreux contre-exemples fleurissent au sein de la littérature. L’ubiquité du gradient latitudinal de diversité et notamment les causes qui lui sont associées, sont fréquemment remises en question. La présente controverse a pour but de réaliser une synthèse des connaissances actuelles sur le gradient latitudinal de biodiversité (GLB). L’objectif n’étant pas seulement d’énoncer les mécanismes et les hypothèses impliqués dans la formation d’un tel patron de distribution mais également de comprendre les exceptions actuelles et passées associées à ce gradient. Enfin, nous nous intéresserons aux méthodologies et aux différents niveaux d’étude employés dans les analyses de la répartition spatiale de la biodiversité.
Problématiques de la controverse
- Quels sont les processus et les mécanismes à l'origine de la formation du gradient latitudinal de biodiversité ?
- Le gradient latitudinal de biodiversité est-il intemporel et universel ?
- Comment peut-on expliquer les exceptions au gradient latitudinal de diversité ?
Publiée il y a plus de 9 ans par Université de Montpellier et J. Morarin.Dernière modification il y a plus de 9 ans.