Vous venez de mettre les pieds dans les tréfonds en construction de ControverSciences.
Origin of the HIV-1 group O epidemic in western lowland gorillas
Origin of the HIV-1 group O epidemic in western lowland gorillas
Mirela D’arc, Ahidjo Ayouba, Amandine Esteban, Gerald H. Learn, Vanina Boué, Florian Liegeois, Lucie Etienne, Nikki Tagg, Fabian H. Leendertz, Christophe Boesch, Nadège F. Madinda, Martha M. Robbins, Maryke Gray, Amandine Cournil, Marcel Ooms, Michael Letko, Viviana A. Simon, Paul M. Sharp, Beatrice H. Hahn, Eric Delaporte, Eitel Mpoudi Ngole, Martine Peeters
Les différentes souches du virus HIV-1 peuvent être divisées en quatre groupes principaux : M, N, O et P. Bien que l'origine des groupes M et N ait déjà été identifiée (franchissement de la barrière inter-espèce de SIVcpz depuis différentes communautés de Chimpanzés du sud du Cameroun vers l'Homme), l'origine des groupes O et P reste encore mal connue. Différentes publications indiquent comme origine possible une transmission de SIVgor depuis le Gorille à l'Homme mais les événements de transmission et leur localisation n'ont jamais été précisément déterminés. Les auteurs s'intéressent donc ici à l'épidémiologie de SIVgor en étudiant sa répartition parmi trois des 4 sous-espèces de gorilles (le Gorille des plaines de l'est, le Gorille des plaines de l'ouest et le Gorille des montagnes), au Cameroun et au Gabon. Par ailleurs, les auteurs s'intéressent au rôle d'un facteur de restriction, APOBEC3G, dans la transmission de SIV entre Chimpanzé et Gorille.
Les auteurs ont réalisé plusieurs prélèvements de matières fécales de plusieurs populations de gorilles au Cameroun et au Gabon, puis les ont testé pour la présence de SIVgor, afin d'étudier sa prévalence dans les populations et la distribution des individus séropositifs.
Ils ont ensuite séquencé les gènes viraux isolés et ont replacé les différentes séquences de SIVgor dans une phylogénie en relation avec des séquences déjà connues de SIVcpz et des différents groupes de HIV-1.
Par ailleurs, afin d'étudier la transmission de SIVcpz du Chimpanzé vers le Gorille, les auteurs ont étudié in vitro la dégradation du facteur de restriction APOBEC3G par la protéine virale Vif de SIVcpz (ou de SIVgor). En effet la dégradation de ce facteur de restriction par la protéine Vif de SIV est nécessaire à la permissivité des cellules de l'hôte à l'infection par SIV.
- SIVgor a été détecté uniquement chez le Gorille des plaines de l'ouest du sud du Cameroun (quelques communautés uniquement). SIVgor serait donc beaucoup moins étendu et prévalent que SIVcpz.
- Deux souches de virus SIVgor (identifiées au sud ouest du Cameroun parmi des populations de gorilles déterminées) sont très proches génétiquement du groupe P de VIH-1 ce qui pointe le Gorille des plaines de l'ouest comme étant vraisemblablement l’espèce à l’origine de la transmission.
- Une troisième souche de SIVgor (identifiée au centre du Cameroun) est très proche génétiquement du groupe O de HIV-1. Le gorille des plaines de l'ouest serait donc ici aussi l’espèce d’origine de la transmission.
- SIVgor serait apparu après transmission inter-espèce d’une seule souche de SIVcpz. La résistance du facteur de restriction APOBEC3G du Gorille à la dégradation par la protéine Vif de SIVcpz pourrait expliquer le fait qu'il n'y ait eu qu'un événement de transmission.
La zone où SIVgor est détecté dans cette étude est très étroite et se restreint à quelques communautés de gorilles. Cependant, depuis plusieurs décennies, la population de gorilles est en déclin (chasse, perte d’habitat, épidémies d’Ebola, pathogénicité possible de SIVgor chez le Gorille) On pourrait donc envisager que SIVgor était auparavant présent dans une zone géographique beaucoup plus étendue (dans les mêmes communautés ou dans d’autres communautés de gorilles qui se sont éteintes). Une transmission inter-espèce vers l’homme a donc pu avoir lieu en dehors de la zone actuelle de répartition de SIVgor.
L'article apporte des données précises sur la localisation du ou des événements de transmission de SIVgor du Gorille vers l'Homme à l'origine de l'apparition des groupes P et O de HIV-1. Ces données peuvent désormais être confrontées aux différentes hypothèses de transmission de SIV (SIVgor ici) du singe (ici Gorille) vers l'Homme : transmission "naturelle" par contact entre Homme et Gorille (chasse...), transmission suite à des campagnes de vaccination, transmission dans le cadre d'expériences de transmission de plasmodiums simiens à l'Homme etc.
Dernière modification il y a plus de 10 ans.
Adaptation de HIV-1 à son hôte humain
Adaptation of HIV-1 to its human host.
Louise V. Wain, Elizabeth Bailes, Frederic Bibollet-Ruche, Julie M. Decker, Brandon F. Keele, Fran Van Heuverswyn, Yingying Li, Jun Takehisa, Eitel Mpoudi Ngole, George M. Shaw, Martine Peeters, Beatrice H. Hahn, and Paul M. Sharp
HIV-1 et 2 sont des virus humains de la famille des lentivirus. Les lentivirus contiennent également de nombreux virus simiens dont des SIV présent chez au moins trente espèces de singes africains. HIV-1 est responsable de la pandémie de SIDA et est divisé en quatre groupes : M, N, O et P. Le groupe M est le plus répandu et est issu de SIVcpz_Ptt_ (présent chez le chimpanzé Pan troglodytes troglodytes) tout comme le groupe N, tandis que les groupes O et P sont issus de SIVgor (présent chez le gorille Gorilla gorilla gorilla).
Cette étude s'intéresse aux mécanismes moléculaires du franchissement de la barrière inter-espèces (passage de SIV chez le singe à HIV chez l'homme) en recherchant des sites de changement génétique associé à la transmission inter-espèces qui pourraient représenter une adaptation de SIVcpz à l'hôte humain.
Trois expériences sont réalisées dans cet article.
La première expérience est une analyse de séquence de 12 souches de SIVcpz_Ptt_ et de 3 souches de HIV-1 à la recherche d'une modification génétique conservée chez tous les SIVcpz_Ptt_ mais pas chez HIV-1.
Dans la seconde expérience, les auteurs comparent des souches de HIV-1 adaptées à ce répliquer chez le chimpanzé avec les souches de HIV-1 "parentales" dont elles sont issues.
Enfin, dans une dernière expérience, l'effet de la mutation trouvée lors de la première expérience sur la réplication du virus est étudié à la fois chez le chimpanzé et l'humain.
- L'acide aminé 30 de la protéine p17 codée par le gène Gag est conservé chez SIVcpz_Ptt_ et chez SIVgor sous forme d'une méthionine (M), mais est muté dans le HIV en un acide aminé basique (arginine R ou lysine K). Donc une forte pression de sélection s'exerce sur cet acide aminé selon l'espèce et suggère un rôle important dans la spécificité à l'hôte. De plus pour chacun des passages du singe à l'homme cette substitution de methionine en arginine ou lysine a eu lieu.
- Dans les souches de HIV-1 adaptées au chimpanzé, l'une des mutations ayant eu lieu est cette substitution de gag30 d'une argine ou lysine en méthionine.
- Concernant la réplication du virus, si gag30 est une méthionine le virus se réplique mieux chez le chimpanzé que chez l'homme tandis que si gag 30 est une lysine le virus se réplique mieux chez l'homme que chez le singe. Donc cette mutation est importante pour une meilleure réplication adaptée à l'hôte.
Cet article généralise la situation à tous les HIV-1 avec seulement 3 souches et pointe pourtant le fait que le sous-type C de HIV-1 M possède une méthionine et non un résidu basique. De plus ce sous-type est très répandu et n'est pas un sous-type ancestral, ce qui suggère qu'il y a eu une réversion de l'arginine en méthionine. Ceci montre que cette substitution n'est pas nécessaire à la propagation du virus chez l'humain, mais reste possiblement un facteur aidant le franchissement de la barrière inter-espèces.
En outre, aucune hypothèse convaincante n'est donnée pour expliquer le fait qu'une mutation d'une protéine de matrice améliore la réplication virale selon l'hôte.
A l'inverse, on peut noter qu'une certaine rigueur a été observée puisque seules les mutations conservées dans les 12 séquences de SIVcpz ont été conservées et que les acides aminés retrouvés chez HIV-1 avec une nature physico-chimique proche de l'acide aminé présents chez SIV n'ont pas été pris en compte.
L'article apporte une nouvelle hypothèse pouvant expliquer le franchissement de la barrière inter-espèces. De plus, il démontre qu'une seule mutation ayant eu lieu après des transfusions sanguines de virus permet d'améliorer la réplication d'un lentivirus selon son hôte.
Dernière modification il y a plus de 10 ans.
Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1
Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1
Brandon F. Keele, Fran Van Heuverswyn, Yingying Li, Elizabeth Bailes, Jun Takehisa, Mario L. Santiago,*, Frederic Bibollet-Ruche, Yalu Chen, Louise V. Wain, Florian Liegeois, Severin Loul, Eitel Mpoudi Ngole, Yanga Bienvenue, Eric Delaporte, John F. Y. Brookfield, Paul M. Sharp, George M. Shaw, Martine Peeters, and Beatrice H. Hahn,
SIVcpz a été jusqu'à présent détecté uniquement chez des Chimpanzés de l’espèce Pan troglodytes troglodytes vivants en captivité. Le but des auteurs est ici d’étudier la prévalence du virus dans des populations de chimpanzés P.t.t sauvages.
L’espèce de Chimpanzés Pan troglodytes est divisée en 4 sous espèces dont Pan troglodytes troglodytes (localisation au sud du Cameroun) et P. t. schweinfurthi_i qui sont les deux espèces connues comme réservoirs de SIVcpz (SIVcpz_Ptt et SIVcpz_Pts_).
Cette étude constitue la première étude épidémiologique de SIVcpz chez des Chimpanzés P.t.t sauvages en Afrique Centrale de l’Ouest.
A partir de prélèvements sur le terrain, les auteurs ont réalisé un test des échantillons avec des anticorps anti HIV-1, puis un séquençage (microsatellites) des échantillons positifs. Ils ont ainsi pu isoler les différentes souches endémiques (des populations de chimpanzés P.t.t locales) de SIVcpz.
Ils ont ensuite déterminé la prévalence de SIVcpz_Ptt_ dans les 10 communautés (10 sites différents) de chimpanzés P.t.t identifiées au Sud du Cameroun.
Ils ont enfin étudié les relations phylogénétiques de ces souches de SIVcpz_Ptt_ avec d’autres souches de SIVcpz et des souches de HIV-1 déjà connues (en utilisant les séquences de pol et env).
Ils ont identifié 5 communautés sans individus positifs, et 5 communautés avec individus positifs (avec des prévalences allant de 4% à 35%).
Ils observent un regroupement phylogéographique des souches (chaque communauté de chimpanzés Ptt isolée géographiquement est associée à une souche de SIVcpz_Ptt_ unique).
Les auteurs ont identifié des souches de SIVcpz_Ptt _présentes dans 3 communautés de Chimpanzés (une au sud-centre et 2 au sud-est du Cameroun) qui sont plus proches génétiquement des groupes M et N de HIV-1 qu’aucune autre souche de SIVcpz isolée jusqu'à présent.
Les auteurs ont ainsi identifié 3 communautés de chimpanzés P.t.t précises (situés dans seulement 2 sites géographiques distincts, deux communautés étant très proches géographiquement) à l’origine des événements de transmission de SIVcpz (SIVcpz_Ptt_ plus précisément donc) à l’Homme, responsables de l’apparition des groupes M et N de HIV-1.
Les auteurs rappellent qu’étant donnée la grande diversité génétique des souches de SIVcpz, et le fait qu’aucun prélèvement n’aie encore été réalisé dans de nombreuses zones d’Afrique centrale et de l’ouest, il est très probable qu’il existent d’autres souches de SIVcpz pas encore découvertes qui ont pu ou pourraient dans le futur franchir la barrière inter-espèce vers l’Homme.
D’autres événements de transmission vers l’Homme on pu se produire et nous sont encore inconnus (pour les groupes M et N de HIV-1 ici mais aussi pour d’autres groupes).
L'article donne une localisation précise et robuste des événements probables de transmission inter-espèce à l’origine de l’apparition des groupes M et N de HIV-1.
Les événements de transmissions sont avec certitude très localisés, et pourront être confrontés avec les différentes hypothèses de transmission.
Dernière modification il y a plus de 10 ans.
Des chasseurs d'Afrique Centrale exposés au virus de l'immunodéficience simienne (SIV)
Central African Hunters Exposed to Simian Immunodeficiency Virus
Marcia L. Kalish, Nathan D. Wolfe, Clement B. Ndongmo, Janet McNicholl, Kenneth E. Robbins, Michael Aidoo, Peter N. Fonjungo, George Alemnji, Clement Zeh, Cyrille F. Djoko, Eitel Mpoudi-Ngole, Donald S. Burke, Thomas M. Folks
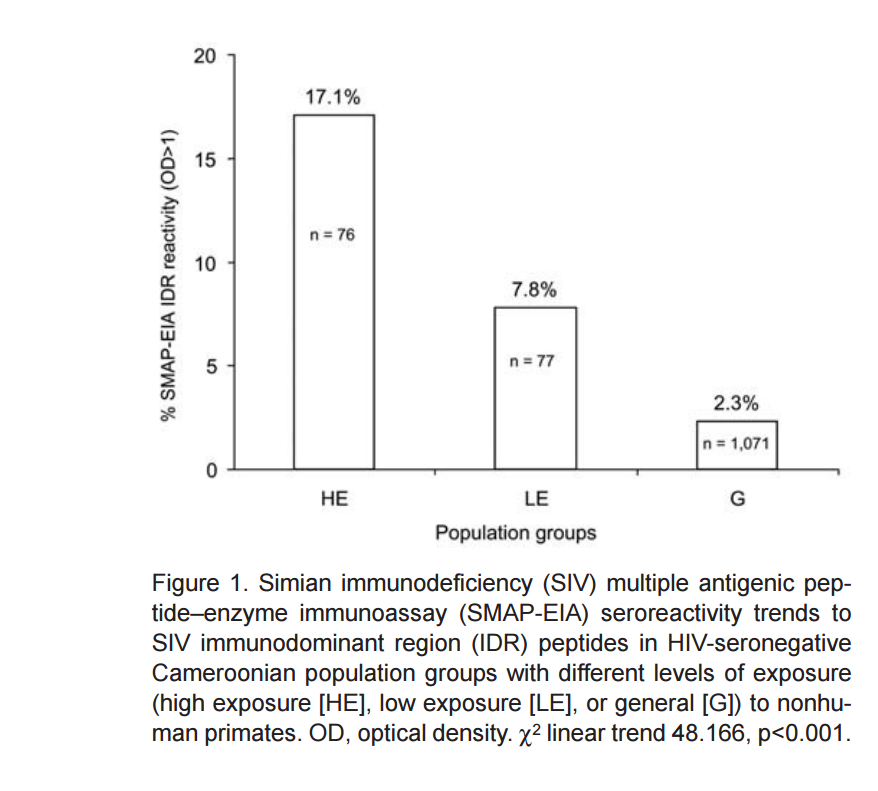
Séroréactivité de groupes fortement exposés, peu exposés, ou généralement exposés, à l'IDR SIV testé par SMAP-EIA
(Kalish 2005)
La phylogénie a montré que HIV-1 et HIV-2 sont apparus chez l'homme à partir de transmissions indépendantes cross-espèces de SIV à partir de primates non humains (NHP): chimpanzés ou sooty mangabeys. La chasse et la consommation de viande de NHP est très répandue en Afrique subsaharienne et expose l'homme à leur sang ou leurs fluides corporels.
Les auteurs ont testé plusieurs échantillons sériques de personnes ayant été exposées à ces fluides, en cherchant à détecter des SIV.
Les auteurs ont mis au point un SMAP-EIA (SIV multiple antigenic peptide-based enzyme immunoassay) qui détecte soit une région immunodominante de gp41 (IDR), soit des anticorps anti-peptide viral V3 proventant de 16 souches différentes de SIV. Les échantillons de plasma proviennent du Cameroun et sont répartis en 3 groupes :
1) fortement exposés : personnes de villages isolés chassant, dépeçant ou ayant des NHP comme animaux de compagnie.
2) peu exposés : personnes des mêmes villages moins exposées aux NHP.
3) contrôle, groupe général : personnes de zones moins isolées, ayant accès à de la viande de NHP, mais peu exposées au sang ou aux fluides.
Les personnes les plus exposées sont les plus séroréactives pour l'IDR SIV testé (Groupe 1) 17.1% ; groupe 2) 7.8% ; groupe 3) 2.3%. (Figure 1) ).
Une personne a présenté une forte réponse humorale au peptide viral V3, mais une très faible réponse cellulaire (faibles résultats par ELISPOT où les cellules sont stimulées par gag). Cela signifie que cette personne a été exposée à un SIV non-viable ou présentant des défauts.
Cet article montre que les personnes chassant et/ou consommant de la viande de brousse issue de primates s'exposent à une infection (parfois productive = pathogénique) par le SIV, car plus de 10 souches semblent déclencher une réponse immunitaire chez l'homme. Ces infections sont souvent restreintes à des zones reculées d'Afriques et ne provoquent donc pas d'épidémie. Cependant, l'article précise que le désenclavage en cours dans ces régions va augmenter la probabilité d'infections productives cross-espèces.
Dernière modification il y a plus de 10 ans.
Origine hybride du SIV chez les chimpanzés
Hybrid Origin of SIV in Chimpanzees
Elizabeth Bailes, Feng Gao, Frederic Bibollet-Ruche, Valerie Courgnaud, Martine Peeters, Preston A. Marx, Beatrice H. Hahn, Paul M. Sharp
Les groupes M et N de HIV-1 sont issus du SIVcpz_Ptt_ présent chez le chimpanzé Pan troglodytes troglodytes. L'origine du SIVcpz est elle-même inconnue et pourrait permettre à la fois de localiser les premiers foyers de SIVcpz et de mieux comprendre la transmission inter-espèces. Dans ce court article, les autres recherchent les origines de SIVcpz.
L'expérience de cet article consiste en une comparaison de 8 espèces de SIV différentes (gsn, syk, sm, rcl, agm, mnd, lhoest et col). Des séquences de gag, pol et env, jugées comme non recombinantes sont comparées avec celles de SIVcpz.
SIVcpz semble être notamment issu d'une recombinaison de SIVrcm (pour le gène pol) présent chez Cercocebus torquatus et SIVgsn (env) présent chez Cercopithecus nictitans deux singes présents au Cameroun et chassés par le chimpanzé.
Dans cet article, seules les protéines de structures virales sont analysées. Il aurait été très intéressant d'analyser les protéines accessoires notamment responsables de la lutte contre les facteurs de restriction.
De plus, tout cet article repose uniquement sur des données phylogénétiques de comparaisons statistiques de séquences génétiques. Les analyses ont été découpées en comparaisons de groupe de quatre espèces de SIV et l'absence d'informations sur les tests statistiques réalisés ne permet pas de savoir si cet enchaînement de comparaisons ne multiplie pas le risque statistique à chaque fois.
Cet article montre qu'un lentivirus peut passer la barrière inter-espèces de façon naturelle par le biais de recombinaisons et que celui-ci peut se répandre dans la nouvelle population cible. De plus, cette transmission semble être le résultat de la chasse de singes par les chimpanzés ce qui met en avant l'hypothèse que la chasse du chimpanzé a pu permettre la transmission du SIVcpz à l'homme en HIV-1.
Enfin, ces informations permettent de faire l'hypothèse que le Cameroun est le lieu de départ de l'épidémie de SIVcpz ce qui concorde avec les hypothèses de transmissions singe-homme.
C'est un article relativement vieux (2003) aujourd'hui et le sujet n'a plus vraiment été abordé depuis.
Dernière modification il y a plus de 10 ans.
Tracing the origin and history of the HIV-2 epidemic
Tracing the origin and history of the HIV-2 epidemic
Philippe Lemey, Oliver G. Pybus, Bin Wang, Nitin K. Saksena, Marco Salemi, and Anne-Mieke Vandamme
La pandémie de HIV observée depuis plusieurs décennies est essentiellement due au groupe M de HIV-1, dont l’origine est établie comme étant des événement de transmission inter-espèce de SIVcpz du Chimpanzé vers l’Homme. Les sous types A et B d’HIV-2 sont également responsables d’une épidémie qui est cependant limitée à l’Afrique de l’Ouest. De plus, bien qu'HIV-2 soit très proche génétiquement de SIVsm (SIV des Sooty Mangabeys), on dispose de très peu d’informations concernant la date et la localisation du ou des évènements de transmissions de SIVsm à l’Homme à l’origine de l’apparition d’HIV-2. L’objectif de l’étude est donc de d’obtenir une estimation de la date de transmission de SIVsm et également d’étudier la démographie de HIV-2 au cours de la seconde moitié du XXème siècle (histoire de l’épidémie).
Les auteurs ont construit la phylogénie (avec une approche de maximum de vraisemblance) de SIVsm et des différentes souches de HIV-2 connues à partir d’alignements de séquences des gènes gag et env.
Ils ont également utilisé la théorie de la coalescence pour étudier l’histoire démographique (évolution du nombre de personnes infectées au cours du temps) du sous type A de HIV-2 dans une région rurale de Guinée-Bissau, qui a auparavant été mise en avant comme un possible noyau de l’épidémie de HIV-2.
Estimation de la date de transmission des deux sous-type de HIV-2 (A et B) du Sooty Mangabeys à l’Homme : première moitié du XXéme siècle. Et plus précisément :
- datation du dernier ancêtre commun des différentes souches de HIV-2 type A : 1940 +/-16
datation du dernier ancêtre commun des différentes souches de HIV-2 type B : 1945 +/- 14
Ces dates constituent donc une limite supérieure pour la transmission inter-espèce du SIV du Sooty Mangabey.datation du dernier ancêtre commun aux sous-type A et B de HIV-2 : 1889 +/-33
Cette date constitue une limite inférieure pour la transmission inter-espèce de SIVsm vers l’Homme.
Les auteurs ont mis en évidence une transition démographique de HIV-2 A entre 1955 et 1970 en Guinée-Bissau avec passage au stade d’épidémie.
Les résultats sur l’histoire de l’épidémiologie de HIV-2 ne prennent en compte que la population rurale de Guinée-Bissau infectée par HIV-2. Il ne faut donc pas extrapoler ces résultats à l’épidémie totale due à HIV-2
Selon les auteurs, à la période de transmission estimée (voir limites supérieures et inférieures) les Sooty Mangabeys avaient une aire de répartition plus étendue que lors de la seconde moitié du siècle (plusieurs sous-espèces éteintes ensuite). Ils soulignent donc le fait que leurs résultats sont en accord avec une transmission naturelle de SIV à l’homme (« natural transfer hypothesis »)
Les auteurs soulignent aussi que le climat de guerre en Guinée-Bissau dans les années 60 pourrait être responsable de changements sociétaux à l’origine de la transition démographique du virus observée (passage d’un virus endémique à une épidémie).
Les auteurs discutent aussi d’un possible rôle des injections de sang contaminé, par des médecins, pendant la guerre d’indépendance (1963-1974) sur la propagation de HIV-2 dans la population en Guinée-Bissau. Cependant, rien n’est dit sur une possible intervention de telles injections sur la transmission de SIVsm du Sooty Mangabey à l’Homme qui a eu lieu dans la première moitié du XXème siècle.
Dernière modification il y a plus de 10 ans.
Transmissions en série à l'homme du SIV par injections non stériles et émergence de l'épidémie de HIV en Afrique
Serial human passage of simian immunodeficiency virus by unsterile injections and the emergence of epidemic human immunodeficiency virus in Africa
P. A. Marx, P. G. Alcabes, E. Drucker
Estimation du nombre de jours d'infection aigue (période à fort titre viral) requis pour m mutations. (Marx et al., 2001)
Les HIV ont émergé en Afrique et dérivent de différents SIV. Il s'agit de comprendre l'émergence quasi simultanée de l'épidémie de deux types de HIV génétiquement distincts à partir de sources simiennes différentes localisées dans des zones géographiques séparées.
Il y a deux hypothèses majeures: les HIV pourraient avoir évolué à partir de cross-transmissions anciennes de SIV à l'homme en restant dans des zones enclavées en Afrique (l'épidémie aurait été déclenchée au 20ème siècle par des mouvements de population), ou des facteurs écologiques favorisant la transformation de SIV en HIV épidémiques auraient émergé au 20ème siècle.
Les auteurs montrent qu'une transmission en série de SIV à l'homme aurait permis l'accumulation de mutations adaptives ayant conduit à l'émergence de HIV épidémiques. Ils se sont basés sur l'augmentation d'injections non stériles observée en Afrique après la Seconde Guerre Mondiale, associée à l'introduction de médicaments injectables.
Les auteurs ont établi une phylogénie liant des SIV et des HIV en tenant compte du caractère épidémique ou non des HIV étudiés, avec un fragment du gène gag et en travaillant par neighbor-joining.
Puis ils ont choisi des sous-types de HIV-1 et -2 faiblement épidémiques, et y ont modélisé des mutations touchant de 20 à 200 nucléotides pour trouver un mécanisme de transition génétique pouvant produire des HIV épidémiques. Des taux de mutation in vivo et des données liées à la prolifération virale ont permis d'évaluer l'accumulation de mutations adaptives par génome de SIV et par personne. Les auteurs ont alors évalué des temps de génération d'un HIV à partir d'un SIV, en fonction de la proportion m (nombre de mutations) du génome SIV devant changer, grâce à un modèle informatique (Table 1).
Les plus fortes proximités SIV/HIV ont lieu entre HIV-1 et SIVcpz, et HIV-2 et SIVsm . Les infections au SIV sont des "dead ends" donc SIV ne peut pas être une zoonose: des mutations adaptives sont nécessaires à son épidémie. Les temps de mutations favorisant la transition SIV/HIV sont longs (Table 1) donc elle ne peut pas s'effectuer pendant la phase chronique de l'infection, car le SI de l'individu détruirait les virus. Elle doit donc avoir concerné plusieurs personnes en même temps. Ainsi, il est très improbable que des souches de HIV aient émergé spontanément à partir d'une infection directe de l'homme au SIV. La transformation rapide de 4 ou 5 anciens SIV en le HIV moderne suggèrent un seul évènement moderne s'étant produit à plusieurs endroits: l'analyse bibliographique indique une forte augmentation (75% des ménages) de la réutilisation de seringues plastiques non stériles entre 1959 et 1964 suite à des campagnes internationales de santé publique.
Une étude très intéressante mais qui reste très bibliographique, où les expériences correspondent uniquement à des modélisations informatiques (certes très rigoureuses), et qui semble un peu détachée de la réalité biologique. Un apport expérimental "de paillasse", même limité, aurait été vraiment intéressant pour pouvoir tester les modèles employés (sur les temps de mutation, par exemple, ou la nature "dead end" des infections au SIV) et aurait rendu l'article beaucoup plus clair et convaincant.
Cet article fournit une hypothèse supplémentaire pouvant expliquer l'émergence soudaine, au milieu du 20ème siècle, de l'épidémie de HIV: la réutilisation massive de seringues plastiques lors de campagnes de santé publique vers 1960 aurait pu être le vecteur d'une infection en série de personnes au SIV, ce qui aurait induit une accumulation simultanée de mutations adaptives de ce virus ayant déclenché la transition SIV-HIV, avec plus ou moins de proximité entre les souches selon le SIV initial.
L'article est globalement très bien documenté et semble rigoureux d'un point de vue mathématique. Le lien avec la bibliographie est bien démontré et intéressant. Cependant, le contenu n'est pas toujours clair, et du coup, parfois moins convaincant (voir paragraphe sur les "dead end SIV infections"). Il est parfois difficile de suivre la démarche expérimentale des auteurs, qui s'appuie énormément sur de la modélisation pas toujours très bien décrite.
Dernière modification il y a plus de 10 ans.
Infection d'un chercheur par le virus de l'immunodéficience simienne (SIV)
Infection of a Laboratory Worker with Simian Immunodeficiency Virus
Rima F. Khabbaz, Walid Heneine, J. Richard George, Bharat Parekh, Thomas Rowe, Toni Woods, William M. Switzer, Harold M. McClure, Michael Murphey-Corb, Thomas M. Folks
Les virus de l'immunodéficience simienne (SIVs) sont des lentivirus morphologiquement similaires et proches biologiquement des virus de l'immunodéficience humaine (HIVs). S'ils infectent naturellement certaines espèces de primates (sooty mangabey, singe vert d'Afrique) sans provoquer d'immunodéficience, ils engendrent en revanche des symptômes proches de ceux causés chez l'homme par le syndrome d'immunodéficience acquise (SIDA) lorsqu'ils sont inoculés expérimentalement à d'autres espèces de primates (macaques). De plus, les SIV issus de mangabeys sauvages et de macaques captifs (SIVMAC) sont particulièrement proches génétiquement du HIV-2.
Ainsi, pour la deuxième fois, un cas d'infection au SIV chez l'homme a été détecté en 1992 chez un chercheur américain présentant des anticorps spécifiques de diverses protéines de SIV et de HIV-2. Cette fois-ci, un SIV humain (SIVHU) a été isolé, confirmant l'infection d'un humain par le SIV.
Entre 1988 et 1992, l'équipe a collecté 20 échantillons de sérum prélevés chez un chercheur américain ayant préalablement été déclaré séropositif pour HIV-2 et SIV. Ils ont été analysés par ELISA spécifiques d'anticorps dirigés contre HIV-1, HIV-2 et SIV, et ont mesuré les titres sériques en anticorps pour étudier leur variation. Des Western Blot révélant des anticorps spécifiques de protéines virales (gp120, gp105, p34) ont également été réalisés.
Puis des échantillons de sang ont été prélevés au chercheur infecté. Les PBMC ont été isolés et mis en culture, et une partie d'entre eux a été déplétée des LT CD8+. La multiplication de SIV dans ces cultures a été évaluée par mesures des concentrations de l'antigène viral p27 gag et d'une reverse transcriptase virale dans le surnageant.
Enfin, des PCR ont été réalisées pour détecter SIVHU, et le séquençage de la région env de SIVHU a permis de la comparer à celle de diverses souches représentatives de SIV et HIV.
Le chercheur testé était séronégatif pour HIV-1 à chaque test, et à partir d'avril 1990, séropositif pour HIV-2 et SIV à chaque test ELISA et Western Blot. L'apparition des anticorps se fait graduellement sur le Western Blot, dénotant une séroconversion. Globalement, le titre sérique en anticorps est passé de 1:400 à 1:3200 en deux ans.
Puis, une culture primaire de PBMCs déplétée en CD8+ a été déclarée positive pour SIV, avec [p27 gag] passant de 0 à 0,9ng/mL en 28 jours de culture. SIVHU a été isolé et inoculé à des PBMCs normaux où il s'est développé aussi, avec des résultats similaires.
Ensuite, la PCR a détecté SIVHU et a montré que son LTR est délété de 40bp, comme chez des SIV conventionnels, ce qui les différencie des HIV.
Enfin, la chaîne env de SIVHU séquencée est particulièrement proche de celle de la souche sur laquelle le chercheur travaillait (SIVB670) avec 90% d'homologie, et moins proche de celles d'autres souches de SIV (76% d'homologie).
Les ELISA ont été effectués uniquement avec un peptide antigénique de HIV-2, et pas avec un peptide de SIV. Même si les anticorps anti-HIV-2 sont cross-réactifs avec SIV d'après les auteurs, il aurait été intéressant de le vérifier.
De plus, le premier Western Blot fait intervenir des échantillons protéiques vieux de trois ans. Des détails sur la conservation des échantillons auraient été souhaitables car il y a une forte probabilité de dégradation des protéines testées au cours d'un laps de temps aussi long.
Enfin, il est également regrettable de ne pas avoir plus de données sur les aspects cliniques de cette infection (SIV chez l'homme). Les auteurs supposent que les symptômes seraient proches de ceux du SIDA causé par HIV-2 chez l'homme. C'est bien sûr cohérent mais on ne peut pas exclure le fait qu'aucune immunodéficience ne se déclare chez l'homme, ce qui serait intéressant. De plus, des hypothèses concrètes sur la façon dont SIV a été inoculé auraient été les bienvenues.
Cet article apporte la preuve qu'une infection au SIV du singe à l'homme est possible, que SIV est capable de se multiplier au sein de cellules humaines, et que ce virus peut être isolé des cellules après infection. Reste cependant à savoir si l'infection est stable (les auteurs mentionnent un précédent cas d'infection au SIV où elle ne l'était pas, les taux de SIV présent chutant sans arrêt) et si elle est pathogénique, ce qui rendrait ces données particulièrement pertinentes par rapport à l'épidémie de HIV.
Dernière modification il y a plus de 10 ans.
Une hypothèse testable pour l'origine des différentes souches du VIH : la transfusion de sang de singes dans le cadre d'expériences sur le paludisme.
AIDS, monkeys and malaria
Charles Gilks
Cette courte lettre publiée dans Nature en 1991 propose une hypothèse testable concernant l'origine des différents VIH (virus de l'immunodéficience humaine). Au contraire, les hypothèses existantes à l'époque sont jugées spéculative par l'auteur.
Le VIH-1 est proche d'une souche de VIS (virus de l'immunodéficience simien) de chimpanzé et le VIH-2 est sans doute un proche parent du VIS que l'on trouve chez les singes mangabeys. Au milieu du XXeme siècle, des scientifiques belges ont transfusé du sang de singes (chimpanzés et mangabeys) à des hommes pour tester la possibilité d'une transmission des parasites du paludisme (plasmodium) spécifique de ces espèces.
En ce basant sur les articles datant de l'époque, l'auteur en arrive au chiffre de 34 personnes ayant reçu des injections de sang frais pris à 17 chimpanzés différents. Du sang de ces 34 personnes a par la suite été transfusé à 33 autres êtres humains. Seuls deux cas de transfusion directe mangabey-homme ont été trouvés mais il y a eu des transfusion indirecte passant par l'intermédiaire de macaques.
Si il y a des données qui attestent de l'existence de ces transfusions, l'ampleur de celles-ci est difficilement estimable étant donné que certaines expériences peuvent n'être consignées dans aucune publication. L'auteur précise que la plupart des expériences documentées prennent place en Europe ou aux États-Unis mais qu'il doit être possible de tester son hypothèse en cherchant dans les archives : les données non-publiées - le devenir des singes utilisés - du matériel biologique connecté à l'un des articles.
Cette "lettre" propose une hypothèse, qui plus est, une hypothèse testable. En revanche, le travail de recherche et de documentation auquel l'auteur appelle n'est ni annoncé, ni entamé.
Plus qu'une contribution, la publication de cet article a été une ouverture publique du débat. Toutes les références de la controverses se positionnent en fait par rapport à cette lettre et à l'hypothèse qu'elle formule.
Remarque subjective : J'ai pris connaissance de l'existence de cette lettre par l'intervention d'un parasitologue travaillant sur le paludisme lors d'une conférence sur les transmissions de rétrovirus simiens. Il est impressionnant de constater qu'on dispose de nombreux cas référencés de transfert du singe vers l'homme pour HTLV (1, 2, 3 et 4) alors qu'on ne trouve aucun virus SIV muté qui puisse fournir une ébauche d'explication concernant l'origine du VIH.
Doit-on supposer que les mécanismes qui sont à l'origine de l'apparition du VIH n'opèrent plus aujourd'hui ?
Dernière modification il y a presque 11 ans.
Des expériences scientifiques sont elles à l'origine du transfert du VIS/VIH du singe à l'homme ?
Si la parenté avec les virus de singes désignés par le sigle VIS (Virus de l'Immunodéficience Simienne) donne une idée précise du phénomène à l'origine de l'émergence du VIH (Virus de l'Immunodéficience Humaine), le mécanisme et les événements ayant permis le transfert de ce virus du singe à l'homme restent inexpliqués.
Publiée il y a presque 11 ans par P. Guille Escuret.En 1991, dans l'article AIDS, monkeys and malaria, un chercheur propose une hypothèse testable impliquant la responsabilité de parasitologues : des chercheurs belges ont étudié la possibilité de transfert vers l'homme des parasites responsables du paludisme chez le singe en effectuant des transfusions sanguines inter-espèces.
L'hypothèse ne semble pas avoir eu un grand écho et le travail de vérification auquel elle appelle reste à faire. En passant en revue les autres hypothèses et les connaissances sur les circonstances du transfert, cette controverse est l'occasion de rechercher s'il existe un faisceau de preuves pouvant asseoir le lien entre le SIDA, les singes et la recherche sur le paludisme.
Dernière modification il y a plus de 10 ans.