Vous venez de mettre les pieds dans les tréfonds en construction de ControverSciences.
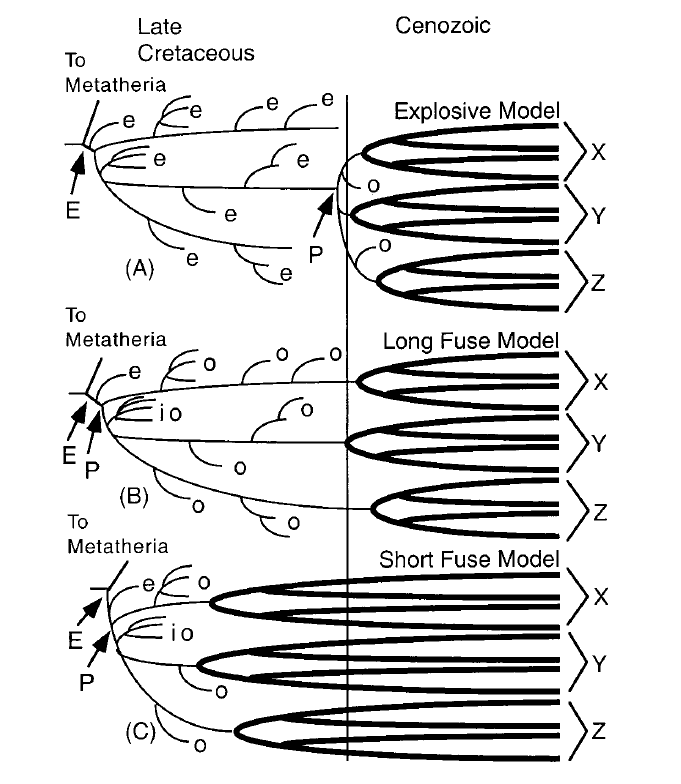
Theories portant sur la diversification des Placentaires. Les lignes en gras représentent des ordres X, Y et Z montrant des diversifications intraordinales à l'intérieur des groupes couronnes. (A) Modèle explosif avec une origine datant du début du Cénozoïque et une diversification du groupe racine puis des groupes couronnes, (B) modèle Long Fuse avec une origine datant du début du Cénozoïque et une diversification des groupes couronnes à la fin du Crétacé, et (C) modèle Short Fuse avec une origine et une diversification des groupes au cours de la fin du Crétacé.
Abbréviations: E, Eutheria; P, Placentalia; e, eutherian stem taxa; o, ordinal stem taxa; io, stem taxa to more than one ordinal crown group
Figure issue de Archibald et Deutschman, 2001
Les premiers vrais mammifères seraient apparus au cours du Trias (-220 Ma), durant le "règne" des dinosaures, et auraient été des animaux de petite taille, nocturnes, insectivores, et possiblement fouisseurs. Il est supposé que la position dominante des dinosaures dans la chaine trophique aurait maintenu les mammifères dans une sorte de « latence évolutive ». Groupe relativement peu diversifié au cours du Mésozoïque, on lui associe à partir de l’extinction de masse du Crétacé - Paléogène (K-Pg; 66 Ma) une myriade de formes et de tailles, retrouvés dans un registre fossile riche.
C’est à partir de ce registre fossile que les premières tentatives de mise en évidence des patrons de diversification ont été faites. George Gaylord Simpson publie en 1944 Tempo and Mode in Evolution, dans lequel il tente de retracer les patrons de diversification des métazoaires. Il y décrit une « explosion » de nouvelles formes de mammifères et d’oiseaux au Paléogène, marquant l’origine des premiers modèles d’évolution.
Les avancées technologiques du XXème siècle ont par la suite permis de séquencer les premiers fragments d'ADN et de mettre en place de nouvelles méthodes pour essayer de dater l’apparition de l’ordre des mammifères placentaires et sa radiation. Les études utilisent alors majoritairement des analyses moléculaires, établissant des phylogénies datées, et recourant peu ou pas à la calibration fossile. Les premières tentatives génèrent des résultats en fort désaccord avec le registre fossile, estimant des dates de divergence bien plus anciennes.
Ainsi, jusqu’il y a un peu plus de 3 ans, trois hypothèses majeures entraient en conflit. Nous devons le nom des modèles présentés ici à Archibald et Deutschman (2001), qui reprennent la littérature de l’époque et tentent d’en tirer une tendance :
• Le modèle explosif (Explosive Model) implique une diversification inter et intra-ordinale des euthériens modernes après la crise K-Pg. L’ancêtre commun des placentaires actuels, selon ce modèle, alterne entre une position post-K-Pg (Hard Explosive Model) ou pré-K-Pg (Soft Explosive Model).
• Le modèle Short Fuse émet quant à lui l’hypothèse que l'origine des placentaires, ainsi que les diversifications inter et intra-ordinales ont toutes eu lieu avant la crise du K-Pg. Les dates de divergence au sein des ordres sont variées, s’étendant jusque dans le Crétacé inférieur.
• Le modèle Long Fuse place les diversifications intra-ordinales après la crise K-Pg, mais postule que la plupart des diversifications inter-ordinales ont eu lieu pendant le Crétacé.
Dans les publications récentes, les patrons proposés sont plus subtils, et globalement, les résultats des méthodes moléculaires se ressemblent. Cependant, il existe encore un conflit majeur d’informations, basé sur l’incohérence de ces résultats avec le registre fossile. Nous présentons ici les différents modèles, ainsi que les techniques ayant mené à leur découverte. De nouveau, nous spécifions l’importance de la distinction entre placentaires (espèces actuelles, leur ancêtre commun et tous ses descendants) et euthériens (placentaires éteints, actuels, et les organismes ayant un ancêtre commun plus récent avec les placentaires qu’avec les métathériens).
I) Les premiers modèles
Le premier modèle, le modèle explosif, est celui soutenu en majorité par les paléontologues. Il doit son nom à la courte période d’explosion de diversité des placentaires, suivant de près la crise du K-Pg. Cette période est considérée courte à l’échelle des temps géologiques, représentant moins de 6% de l’histoire évolutive des euthériens. Malgré la longévité de ce modèle, il n’a toujours pas été réfuté de manière certaine. Des publications récentes, portant sur l’étude de caractères morphologiques (Halliday et al., 2017, Hard Explosive Model) et moléculaires (Phillips et Fruciano, 2018, Soft Explosive Model), continuent de soutenir l’idée d’une diversification des placentaires actuels s’étant effectuée majoritairement au Paléogène.
Le modèle Short Fuse voit le jour suite aux premiers résultats obtenus par inférences phylogénétiques des âges de divergence, grâce à des modèles d’évolution des séquences nucléotidiques ou protéiques, se basant sur des hypothèses d’horloge moléculaire (les mutations génétiques s’accumulent de manière constante dans le génome). Ces premières études placent les diversifications au sein des placentaires modernes avant la crise du K-Pg, allant jusque 90 Ma pour les 1er divergences au sein des lagomorphes (Kumar et Hedges, 1998). Cependant, il a été rapidement montré que les méthodes présentaient de nombreux défauts :
1) Les taux d’évolution moléculaire ne sont pas constants, ni au cours du temps, ni entre les différents clades. Des modèles plus complexes sont donc nécessaires, pour se rapprocher de la réalité.
2) Il est nécessaire de calibrer les arbres dans le temps, en assignant des bornes maximales et minimales à certains nœuds, en fonction de fossiles non controversés, datés, et assez numéraires.
3) Il est nécessaire d’inclure suffisamment de données, pour ne pas créer un biais d’échantillonnage trop important en n’incluant que certains taxons.
Les problèmes méthodologiques de ces premières analyses ont conduit à de très fortes surestimations des âges de divergence, avec des ancêtres supposés prédatant le plus ancien représentant fossile d’un clade. Le décalage entre ces deux estimations temporelles correspond, pour certaines études, à des dizaines de millions d’années, et représente des lignées "fantômes".
Le modèle Long Fuse apparait avec les premières corrections des défauts méthodologiques (Springer et al., 2003), en particulier l’ajout de calibrations fossiles plus nombreuses et mieux réfléchies. L’amélioration des modèles d’évolution des séquences, ayant un degré supérieur de réalisme (taux variables au cours du temps et entre clades) ont aussi permis d’obtenir des phylogénies sans doutes plus proches de la réalité, puisque les résultats obtenus se retrouvent plus en accord avec le registre fossile, notamment concernant les diversifications intra-ordinales. Un consensus émerge dans les études moléculaires récentes : les superordres modernes (Xenarthra, Afrotheria, Laurasiatheria et Euarchontoglires) seraient tous apparus au Crétacé, et les ordres majeurs auraient déjà été présents avant la crise du K-Pg (Meredith et al., 2011; Dos Reis et al., 2014; Tarver et al., 2016). Les dates exactes de divergence et la topologie de l'arbre sont assez variables selon les études en question, mais dépendent fortement de la méthodologie.
II) Etat de l'art
Le modèle Short Fuse a aujourd’hui été mis de côté, au profit des modèles Long Fuse ou Soft Explosive. Mis à part les études morphologiques, qui continuent à placer l’ancêtre placentaire après la crise du K-Pg (O’Leary et al., 2013 ; Halliday et al., 2017), les modèles actuels s’accordent sur deux points :
1) Ils placent l’origine des placentaires modernes au Crétacé.
2) Ils placent la majeure partie des diversifications intra-ordinales après la crise du K-Pg.
Le désaccord principal concerne le timing des diversifications inter-ordinales, qui débutent avant la crise pour le modèle Long Fuse (Tarver et al., 2016 ; Liu et al., 2017), et après la crise pour le modèle Soft Explosive (Phillips et Fruciano, 2018). Pour contribuer à l’estimation des temps de divergence des placentaires, certaines études se sont basées sur des taxons dont la diversification est corrélée à celle des mammifères, comme par exemple les angiospermes (Meredith et al, 2011), ou les exo-parasites de mammifères (Johnson et al., 2018).
D’après Beck et Baillie (2018), le conflit entre les estimations basées sur les données moléculaires et morphologiques serait réglé si le registre fossile était complété, ce qui laisse à penser que la découverte de fossile dans le futur permettrait de mettre fin à la controverse.
III) Difficultés, causes méthodologiques de la controverse
À l’heure actuelle, aucun euthérien datant du Crétacé n’a été assigné de manière certaine aux placentaires (groupe couronne). Une hypothèse avancée dans les années 2000 stipule qu’une évolution moléculaire latente aurait lieu avant que les modifications phénotypiques détectables sur les fossiles n'apparaissent. Ces placentaires du Crétacé manquant au registre fossile ont donc peut-être déjà été trouvés, sans avoir été identifiés, car ces évolutions moléculaires auraient agi à des niveaux physiologiques, ou sur des tissus mous, indétectables à l’état fossile. De plus, l’utilisation des caractères morphologiques présente des défauts non négligeables :
- Les études en génétique du développement montrent que certains caractères morphologiques évoluent de manière codépendante, sous l’influence des mêmes gènes (nombre de cuspides, forme de la dent). Ainsi, lorsque l’on construit des phylogénies, qui considèrent les caractères comme indépendants, on néglige fortement l’une des conditions d’applications.
- En 2007, Springer a mis en évidence un autre phénomène problématique : dans des analyses en « total evidence », alliant caractères morphologiques et moléculaires, lorsque l’on supprime les caractères moléculaires d’espèces actuelles afin de les considérer comme des fossiles, les analyses ne les replacent pas dans leur taxon « réel ». Comment être sûr que les fossiles utilisés pour les calibrations, assignés à certains taxons en fonction de leurs caractères morphologiques, appartiennent en réalité à ces clades?
Selon Phillips et Fruciano (2016, 2018), les taxons présentant des grandes tailles et des longévités élevées sont à l’origine de l’existence de lignées fantômes (fossiles du groupe plus récents que la date de divergence estimée). Ils proposent une correction au niveau des calibrations qui crée, selon Springer et al., (2017), un autre problème : l'apparition de lignées "zombies", avec des fossiles identifiés qui prédatent le moment estimé de la divergence du clade.
IV) Conclusion et ouverture
Il a longtemps été admis que l’extinction des dinosaures non aviaires, à la fin du Crétacé, aurait permis une radiation explosive des mammifères, par libération de niches écologiques. Il est indéniable que l’extinction soudaine d’un grand nombre d’organismes ait pu permettre à d’autres de se diversifier. Cependant, le degré du lien de causalité entre les deux évènements est remis en question par les analyses moléculaires, qui montrent pour la plupart que les ancêtres des placentaires actuels seraient apparus au cours du Crétacé, aux alentours de 80 Ma, et que les premiers ordres auraient vu le jour bien avant l’extinction de masse. Ceci voudrait dire que les ancêtres des placentaires actuels auraient cohabité avec les dinosaures. De nombreux projets de recherche récents semblent indiquer qu’un déclin notable de la diversité spécifique ait été en cours chez les grands dinosaures herbivores, bien avant l’extinction de masse engendrée par la chute de l’astéroïde. Ce déclin, initié au cours du Crétacé, pourrait soutenir l’idée d’une évolution du groupe couronne placentaire plus précoce que ce qui était initialement pensé.
Malgré le charme de cette idée, la controverse reste animée, pour une raison majeure : si de très nombreuses études phylogénétiques placent de manière de plus en plus certaine l’origine et les premières diversifications inter-ordinales des placentaires actuels au cours du Crétacé, on n’en retrouve pas la trace dans le registre fossile. Les paléontologues ne sont pas en mesure d’attribuer de manière certaine les euthériens du Crétacé au groupe couronne des placentaires. Ceci ne permet donc pas de calibrer dans le temps les phylogénies avec certitude, puisque les fossiles trouvés ne peuvent pas être assignés de manière certaine à l’un des nœuds de l’arbre calibré dans le temps des mammifères actuels. En effet, malgré des études morphologiques de plus en plus poussées, et l’utilisation de matrices de caractères alliant des organismes éteints et actuels, la position de ces groupes basaux ou latéraux demeure cryptique, notamment à cause des problèmes de convergence ou d'évolution corrélée des caractères morphologiques. Il est évident que l'utilisation des fossiles est indispensable, car ce sont des témoins directs de l'évolution des organismes. Néanmoins, il faudra allier de nouveaux spécimens et une méthodologie corrigée afin d'espérer résoudre le conflit entre les différents scénarios de diversification.
Publiée il y a plus de 7 ans par A. Stahl et collaborateurs..Dernière modification il y a plus de 6 ans.
Cette synthèse se base sur 12 références.
- • Analyse quantitative de la datation de l'origine et de la diversification des ordres placentaires existants
- • Résoudre les relations entre les mammifères placentaires du Paléocène.
- • Le modèle explosif "souple" de l'évolution des mammifères placentaires
- • Une échelle moléculaire pour l'évolution des vertébrés
- • Impacts de la révolution terrestre du Crétacé et de l'extinction du Crétacé-Paléogène sur la diversification des mammifères
- • Ni les données phylogénomiques, ni les données paléontologiques ne soutiennent une origine des mammifères placentaires au Paléogène.
- • Les relations entre mammifères placentaires et les limites de l'inférence phylogénétique
- • L'ancêtre des mammifères placentaires et leur radiation post crise du Crétacé-Paléogène
- • Des preuves génomiques révèlent que la diversification des mammifères placentaires est ininterrompu par la crise C/T
- • La radiation simultanée des poux d'oiseaux et de mammifères après la limite K-Pg
- • Des améliorations du registre fossile pourraient largement résoudre le conflit entre les estimations morphologiques et moléculaires de la phylogénie des mammifères.
- • Réveiller les morts-vivants : les implications d'un modèle explosif atténué pour le timing de diversification des mammifères placentaires.
Des améliorations du registre fossile pourraient largement résoudre le conflit entre les estimations morphologiques et moléculaires de la phylogénie des mammifères.
Improvements in the fossil record may largely resolve the conflict between morphological and molecular estimates of mammal phylogeny
Robin M. D. Beck, Charles Baillie
Le modèle explosif "souple" de l'évolution des mammifères placentaires
The soft explosive model of placental mammal evolution
Matthew J. Phillips and Carmelo Fruciano
La radiation simultanée des poux d'oiseaux et de mammifères après la limite K-Pg
Simultaneous radiation of bird and mammal lice following the K-Pg boundary
Kevin P. Johnson, Nam-phuong Nguyen, Andrew D. Sweet, Bret M. Boyd, Tandy Warnow, Julie M. Allen
Résoudre les relations entre les mammifères placentaires du Paléocène.
Resolving the relationships of Paleocene placental mammals
Thomas J. D. Halliday, Paul Upchurch and Anjali Goswami
Réveiller les morts-vivants : les implications d'un modèle explosif atténué pour le timing de diversification des mammifères placentaires.
Waking the undead: Implications of a soft explosive model for the timing of placental mammal diversification
Mark S. Springer, Christopher A. Emerling, Robert W. Meredith, Jan E. Janečka, Eduardo Eizirik, William J. Murphy
Drapeaux rouges phylogénomiques: erreurs d'homologie et lignées de zombies dans la diversification évolutive des mammifères placentaires
Phylogenomic red flags: Homology errors and zombie lineages in the evolutionary diversification of placental mammals
John Gatesy, Mark S. Springer
Des preuves génomiques révèlent que la diversification des mammifères placentaires est ininterrompu par la crise C/T
Genomic evidence reveals a radiation of placental mammals uninterrupted by the KPg boundary
Liang Liu, Jin Zhang, Frank E. Rheindt, Fumin Lei, Yanhua Qu, Yu Wang, Yu Zhang, Corwin Sullivan, Wenhui Nie, Jinhuan Wang, Fengtang Yang, Jinping Chen, Scott V. Edwards, Jin Meng, Shaoyuan Wu
Les relations entre mammifères placentaires et les limites de l'inférence phylogénétique
The interrelationships of placental mammals and the limits of phylogenetic inference
James E. Tarver, Mario dos Reis, Siavash Mirarab, Raymond J. Moran, Sean Parker, Joseph E. O’Reilly, Benjamin L. King, Mary J. O’Connell, Robert J. Asher, Tandy Warnow, Kevin J. Peterson, Philip C.J. Donoghue, Davide Pisani
Folie mammalienne : La phylogénie des mammifères n'est-elle pas encore résolue?
Mammal madness: is the mammal tree of life not yet resolved?
Nicole M. Foley, Mark S. Springer and Emma C. Teeling
Datation géomoléculaire et origine des mammifères placentaires.
Geomolecular Dating and the Origin of Placental Mammals
Matthew. J. Phillips
Ni les données phylogénomiques, ni les données paléontologiques ne soutiennent une origine des mammifères placentaires au Paléogène.
Neither phylogenomic nor palaeontological data support a Palaeogene origin of placental mammals
Mario dos Reis, Philip C. J. Donoghue, Ziheng Yang
L'ancêtre des mammifères placentaires et leur radiation post crise du Crétacé-Paléogène
The placental mammal ancestor and the post-K-Pg radiation of placentals
Maureen A. O'Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible, Andrea L. Cirranello
Analyse comparative des patrons de diversification des mammifères
A Comparative Study of Mammalian Diversification Pattern
Wenhua Yu, Junxiao Xu, Yi Wu, Guang Yang
Des jeux de données phylogénomiques fournissent des précisions sur l'estimation de l'échelle de temps de la phylogénie des mammifères placentaires
Phylogenomic datasets provide both precision and accuracy in estimating the timescale of placental mammal phylogeny
Mario dos Reis, Jun Inoue, Masami Hasegawa, Robert J. Asher, Philip C. J. Donoghue and Ziheng Yang
Impacts de la révolution terrestre du Crétacé et de l'extinction du Crétacé-Paléogène sur la diversification des mammifères
Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification
Robert W. Meredith, Jan E. Janecka, John Gatesy, Oliver A. Ryder, Colleen A. Fisher, Emma C. Teeling, Alisha Goodbla, Eduardo Eizirik, Taiz L. L. Simão, Tanja Stadler, Daniel L. Rabosky, Rodney L. Honeycutt, John J. Flynn, Colleen M. Ingram, Cynthia Steiner, Tiffani L. Williams, Terence J. Robinson, Angela Burk-Herrick, Michael Westerman, Nadia A. Ayoub, Mark S. Springer, William J. Murphy,
L'adéquation de la morphologie pour reconstruire l'histoire de l'origine des mammifères placentaires
The Adequacy of Morphology for Reconstructing the Early History of Placental Mammals
Springer, Burk-Herrick, Meredith, Eizirik, Teeling, O'Brien, Murphys
La montée retardée des mammifères actuels
The delayed rise of present-day mammals.
Olaf R. P. Bininda-Emonds, Marcel Cardillo, Kate E. Jones, Ross D. E. MacPhee, Robin M. D. Beck, Richard Grenyer, Samantha A. Price, Rutger A. Vos, John L. Gittleman & Andy Purvis
La montée de l'oxygène ces 205 derniers millions d'années et l'évolution des grands mammifères placentaires
The Rise of Oxygen over the Past 205 Million Years and the Evolution of Large Placental Mammals
Paul G. Falkowski, Miriam E. Katz, Allen J. Milligan, Katja Fennel, Benjamin S. Cramer, Marie Pierre Aubry, Robert A. Berner, Michael J. Novacek, Warren M. Zapol
La diversification des mammifères placentaires et la limite du Crétacé-Tertiaire
Placental mammal diversification and the Cretaceous-Tertiary boundary
Mark S. Springer, William J. Murphy, Eduardo Eizirik, and Stephen J. O’Brien
Analyse quantitative de la datation de l'origine et de la diversification des ordres placentaires existants
Quantitative Analysis of the Timing of the Origin and Diversification of Extant Placental Orders
J. David Archibald, Douglas H. Deutschman
Preuves moléculaires de la divergence précoce des mammifères placentaires
Molecular evidence for the early divergence of placental mammals
Simon Easteal
Une échelle moléculaire pour l'évolution des vertébrés
A molecular timescale for vertebrate evolution
Sudhir Kumar, S. Blair Hedges
Explosions évolutionnaires et la mèche phylogénétique
Evolutionary explosions and the phylogenetic fuse
Alan Cooper, Richard Fortey
"À quel moment se place la diversification des mammifères placentaires actuels par rapport à la crise du Crétacé-Paléogène? "
Le groupe des mammifères est un clade monophylétique que l’on peut subdiviser en trois taxons actuels : les monotrèmes, les marsupiaux, et les placentaires. Les premiers mammifères, que l’on peut distinguer de manière non controversée des reptiles mammaliens, seraient apparus aux alentours de 220 Ma, au Trias. Si, aujourd’hui, le nombre de mammifères est estimé à plus de 5400 espèces, leur diversité était assez restreinte jusque la fin du Crétacé (66 Ma). De par sa très grande disparité, les patrons de divergence de ce groupe ont été étudiés sous de très nombreux angles. Pourtant, à l’heure actuelle, il n’existe toujours pas de consensus quant au timing des grands évènements de diversification ayant donnés lieu à la richesse des mammifères placentaires existants.
Dans un premier temps, il est important de mettre au clair une notion importante de cette controverse : ici, le débat concerne seulement la diversification des ordres placentaires actuels. Il faut bien faire la différence entre
Le débat existe uniquement concernant le groupe couronne des placentaires, par conflit entre informations morphologiques provenant de fossiles, et d’informations moléculaires obtenues à partir des séquences nucléotidiques et protéiques. Si quelques euthériens basaux sont connus du Crétacé, jusqu'à maintenant, aucun fossile de cette époque n’a été attribué de manière certaine au groupe couronne des placentaires (Halliday et al., 2017). Ainsi, les études se basant sur des caractères morphologiques placent les principales diversifications après la 5ème crise d'extinction de masse (O' Leary et al., 2013). Ceci est assez problématique, car la majorité des analyses phylogénétiques à base de caractères moléculaires indiquent un ancêtre commun ayant émergé au Crétacé (Tarver et al., 2016; Liu et al., 2017; Phillips et Fruciano, 2018). Ainsi, le patron exact de diversification de ce groupe reste une énigme, que cela concerne l’origination, la divergence des différents ordres, ainsi que la diversification au sein de ces différents ordres.
Publiée il y a plus de 7 ans par I. Voet.Dernière modification il y a plus de 7 ans.